13.3. Основные пути и способы видообразования
В территориальном аспекте в пространстве новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида. Такое видообразование называется аллопатрическим (от греч. alios — иной, patris — родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. sym — вместе, patris — родина). В филогенетическом аспекте (во времени) новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим.
Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов — гибридогенное видообразование. Кратко опишем эти основные пути видообразования.
Аллопатрическое видообразование (называемое иногда географическим) иллюстрируется приведенными выше примерами возникновения вида у больших чаек и в группе австралийских мухоловок. При аллопатрическом видообразовании новые виды могут возникнуть путем фрагментации, распадения ареала широко распространенного родительского вида. Примером такого процесса может быть возникновение видов ландыша (см. гл. 6). Другой способ аллопатрического видообразования — видообразование при расселении исходного вида, в процессе которого все более удаленные от центра расселения периферийные популяции и их группы, интенсивно преобразуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, амфибий, насекомых.
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
Симпатрическое видообразование. При видообразовании симпатрическим путем новый вид возникает внутри ареала исходного вида.
Первый способ симпатрического видообразования — возникновение новых видов при быстром изменении кариотипа, например при автополиплоидии. Известны группы близких видов (обычно растений) с кратными числами хромосом (см. рис. 6.28). Так, например, в роде хризантем (Chrysanthemum) все формы имеют число хромосом, кратное 9, 18, 27, 36, 45, ..., 90. В родах табака (Nicotiana) и картофеля (Solanum) основное, исходное, число хромосом равно 12, но имеются формы с 24, 48, 72 хромосомами. В таких случаях можно предположить, что видообразование шло путем автополиплоидии — посредством удвоения, утроения, учетверения и т. д. основного набора хромосом предковых видов. Процессы полиплоидизации хорошо воспроизводятся в эксперименте посредством задержки расхождения хромосом в мейозе в результате воздействия, например, колхицином. Известно, что полиплоиды могут возникать и в природных условиях. Возникшие полиплоидные особи могут давать жизнеспособное потомство лишь при скрещивании с особями, несущими то же число хромосом (или при самоопылении). В течение немногих поколений в том случае, если полиплоидные формы успешно проходят «контроль» естественного отбора и оказываются лучше исходной диплоидной, они могут распространиться и сосуществовать совместно с породившим их видом (рис. 13.4) или, что бывает чаще, просто вытеснить его.
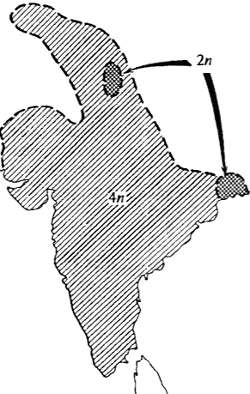
Рис. 13.4. Пример возникновения репродуктивной изоляции при полиплоидизации: обитающий по всему Индостану тетраплоидный вид растений Dicanthium annulatum, несомненно, возник из диплоидной предковой формы, ныне занимающей небольшой дизъюнктный ареал (по Н. Россу, 1962)
Полиплоидные формы, как правило, крупнее и способны существовать в более суровых физико-географических условиях. Именно поэтому в высокогорьях и в Арктике число полиплоидных видов растений резко увеличено (рис. 13.5). Среди животных полиплоидия при видообразовании играет несравненно меньшую роль, чем у растений, и во всех случаях связана с партеногенетическим способом размножения (например, у иглокожих, членистоногих, аннелид и других беспозвоночных).
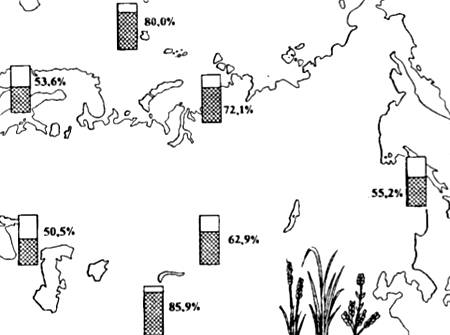
Рис. 13.5. Распространение полиплоидных видов цветковых (в процентах к общему числу видов флоры) в разных частях Евразии (по данным разных авторов из Н.В. Тимофеева-Ресовского и др., 1977)
Второй способ симпатрического видообразования — путем гибридизации с последующим удвоением числа хромосом — аллополиплоидия (см. ниже).
Наконец, последним, достаточно изученным способом симпатрического видообразования является возникновение новых форм в результате сезонной изоляции. Известно существование ярко выраженных сезонных рас у растений, например у погремка Alectorolophus major (см. гл. 10), раннецветущие и позднецветущие формы которого полностью репродуктивно изолированы друг от друга, и, если условие отбора сохраняется, лишь вопрос времени — когда эти формы приобретут ранг новых видов. Аналогично положение с яровыми и озимыми расами проходных рыб; возможно, что эти формы уже являются разными видами, очень схожими морфологически, но изолированными генетически (виды-двойники).
Особенность симпатрического пути видообразования — возникновение новых видов, морфофизиологически близких к исходному виду. Так, при полиплоидии увеличиваются размеры, но общий облик растений сохраняется, как правило, неизменным; при хромосомных перестройках наблюдается та же картина; при экологической (сезонной) изоляции возникающие формы также обычно оказываются морфологически слабо различимыми. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждого из родительских видов (но также имеющая признаки, характерные для двух исходных видов).
Филетическое видообразование. При филетическом видообразовании вид, весь в целом изменяясь в чреде поколений, превращается в новый вид, который можно выделить, сопоставляя морфологические характеристики этих групп.
Филетическое видообразование включает стазигенез — развитие вида во времени с постепенным изменением одной и той же экологической ниши, и анагенез — развитие вида с приобретением каких-то новых принципиальных приспособлений, позволяющих ему образовать совершенно новую, более широкую экологическую нишу. Примером стазигенеза может быть развитие основного ствола верхнеплиоценовых моллюсков рода Giraulus (см. рис. 6.5).
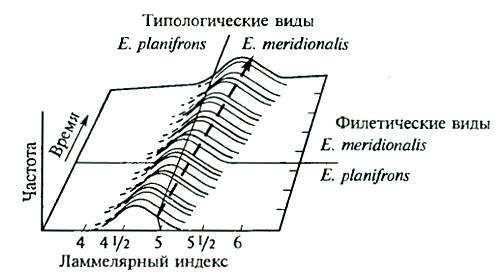
Рис. 13.6. Пример филетического видообразования в ряду ископаемых европейских слонов (Elephas planifrons — Е. meridionalis) по ламеллярному индексу (количеству эмали на зубах) (из В. Гранта, 1980)
Ясно, что в этом случае возможно лишь сопоставление морфологических характеристик, так как изучать результаты филетической эволюции можно лишь с привлечением палеонтологического материала (рис. 13.6). При этом всегда остается возможность того, что на каком-то этапе эволюции от единого филетического ствола могли дивергировать другие группы и филетическое видообразование могло оказаться на самом деле дивергентным. Поэтому в «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одном из отрезков жизни вида (фратрии).
Заметим, что границы между отдельными видами в филетическом ряду форм провести невозможно — она всегда будет условной (см. гл. 12).
Дивергентное видообразование (кладогенез). Ч. Дарвин считал этот тип видообразования самым распространенным (единственный рисунок в «Происхождении видов» посвящен именно этому типу видообразования). Примерами этого типа видообразования — возникновение новых видов в результате разделения единой предковой формы — являются возникновение нескольких видов сигов вокруг Ирландского моря (см. рис. 6.10) и дивергенция дарвиновых вьюрков на Галапагосах (см. рис. 6.12) и эволюция североамериканских дрозофил группы pseudoobscura — persimilis (см. рис. 6.27).
Гибридогенное видообразование (синтезогенез или сингенез). Этот тип видообразования обычен у растений: по некоторым подсчетам, более 50% видов растений представляют собой гибридогенные формы — аллополиплоиды. Укажем лишь на некоторые. Культурная слива (Prunus domestica) с 2n = 48 возникла путем гибридизации терна (P. spinosa, 2n = 32) с алычой (P. divaricata, 2n= 16) с последующим удвоением числа хромосом. Некоторые виды пикульника, малины, табака, брюквы, полыни, ириса и других растений — такие же аллополиплоиды гибридогенного происхождения.
Интересен случай возникновения нового симпатрического вида у Spartina townsendii (2л = 120) на основе гибридизации с последующим удвоением числа хромосом местного английского вида S. stricta (2n = 50) и завезенной в 70-х годах XIX в. из Северной Америки S. alternifolia (2n = 70). Сейчас ареал этого вида интенсивно расширяется за счет сокращения ареала местного европейского вида. Другой пример гибридогенного вида, возникшего симпатрически,— рябинокизильник (Sorbocotaneaster), сочетающий признаки рябины и кизильника и распространенный в середине 50-х годов в лесах южной Якутии по берегам среднего течения р. Алдан (К.М. Завадский). В результате гибридогенного видообразования особенно часто могут образовываться комплексы видов (или так называемых полувидов), связанных между собой гибридизацией,— сингамеоны (В. Грант). В случае таких гибридных комплексов иногда бывает трудно обнаружить четкие границы между отдельными видами, хотя виды как устойчивые генетические системы выделяются вполне определенно.
Все четыре основные формы видообразования во времени схематично показаны на рис. 13.7.
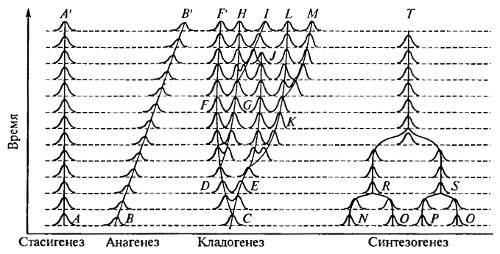
Рис. 13.7. Основные формы филетического (во времени) видообразования (из Н.Н. Воронцова, 2001)
