13.2. Примеры видообразования
В настоящее время изучены различные пути возникновения новых видов в процессе микроэволюции.
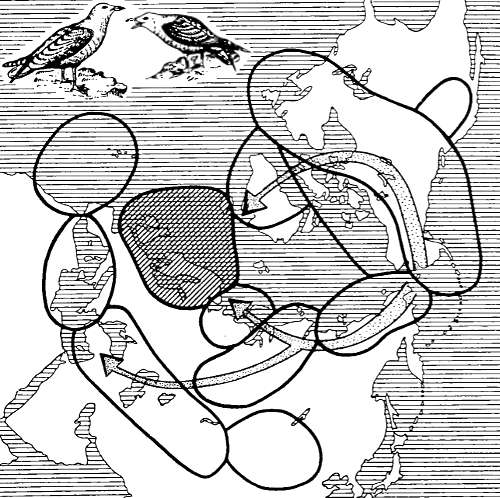
Рис. 13.2. Цепь подвидов больших чаек: серебристая — клуша-хохотунья (Larus argentatus, L. fuscus) (по Н.В. Тимофееву-Ресовскому и др., 1977)
Видообразование в цепи подвидов больших чаек. По побережьям Балтийского и Северного морей живут не скрещиваясь два вида крупных чаек: серебристая чайка (Larus argentatus) и клуша-хохотунья (L. fuscus). Эти два вида объединяются друг с другом через непрерывную цепь подвидов, охватывающих Северную Евразию, с одной стороны, и Гренландию и Северную Америку — с другой (рис. 13.2). Несколько сотен тысяч лет назад в районе современного Берингова пролива обитала предковая форма этих чаек. Впоследствии они (биологически связанные либо с побережьями морей, либо с крупными внутриконтинентальными водоемами) начали распространяться на восток и запад, образовав к нашему времени две непрерывные цепи подвидов. Особи всех соседних подвидов скрещиваются в природе и дают плодовитое потомство. В районе же Северного и Балтийского морей произошла встреча конечных звеньев восточной и западной цепи подвидов. Накопленные в процессе микроэволюции отдельных подвидов различия в биологии (особенности образа жизни, некоторые морфологические особенности и др.) оказались достаточными для возникновения двух новых видов. Если по каким-либо причинам непрерывная цепь подвидов где-нибудь разорвется, то возникнут два четко различающихся вида. Сейчас же видообразование в этой группе чаек находится как бы «в процессе становления» (in statu nascendi), давая нам возможность наблюдать эволюцию в действии. Сходный пример цепи подвидов, крайние звенья которой могут быть названы видами, описан для грауса и белой куропатки (см. 12.3).
Видообразование в группе австралийских мухоловок. В настоящее время в Австралии распространены два близких вида мухоловок рода Pachycephala (P. rufogularis и P. inornata). В результате тщательного анализа палеогеографии и палеоклиматологии удалось восстановить возможный путь образования такой странной на первый взгляд ситуации, когда один из близких видов обитает лишь на небольшом пространстве, занимаемом другим видом (рис. 13.3). Вначале существовал лишь один предковый вид, имевший широкий ареал (рис. 13.3, 1). В связи с наступлением засушливого периода, изменениями в растительности и смещениями дождевых зон возникла и развилась значительная изоляция между западной и восточной частями видового населения (рис. 13.3, 2, 3). Существуя в несколько иных условиях и при отсутствии нивелировок достигаемых различий (при отсутствии скрещивания), две формы приобрели видовую самостоятельность (рис. 13.3, 4). Затем вновь получила распространение сравнительно влаголюбивая растительность и западный вид смог расселиться на восток, где он встретился с родственным видом, приобретшим какие-то специфические особенности. Различия между этими прежде генетически близкими формами оказались настолько значительны, что определили возможность их самостоятельного существования без скрещивания. В сравнительно недавнее время в результате нового иссушения климата вновь возникла изоляция между западной и восточной частями населения одного из видов (рис. 13.3, 5).
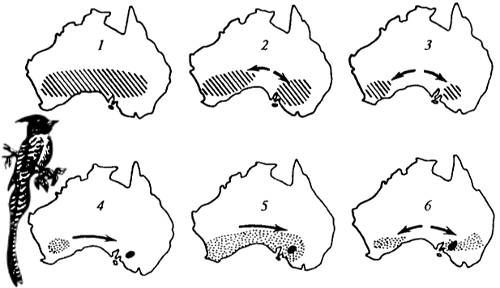
Рис. 13.3. Видообразование в группе австралийских мухоловок Pachycephala (из Э. Майра, 1968) Современное распространение показано на карте 6. Расширения и сокращения ареалов связаны с изменениями в растительности, вызванными, в свою очередь, историческими смещениями дождевых зон в разные периоды эволюции
