20.3. Монофилия и полифилия. Сетчатая эволюция
Проблема монофилии — пример того, как накопление новых данных заставляет ограничить сферу приложимости некоторых, еще недавно казавшихся всеобщими принципов.
Принцип монофилии — происхождения групп от одной предковой формы — основа классической филогенетической систематики. Однако в современной трактовке принципа монофилии произошли изменения. Еще Ч. Дарвин считал, что новый вид происходит не от одной, а от многих особей. Сегодня это положение полностью подтверждено развитием учения о микроэволюции: ясно, что элементарная эволюционная единица не особь, а отдельная популяция, поэтому выражение «происхождение от единого предка» нельзя понимать буквально, в смысле происхождение от единой особи. Монофилетической надо считать группу, происходящая от одной группы того же таксономического ранга. Например, семейство современных ластоногих (ушастые и настоящие тюлени) произошло от групп видов, принадлежащих, видимо, даже к разным семействам отряда хищных. Однако это не дает основания для признания современного отряда ластоногих полифилетической группой: весь отряд ластоногих произошел от одного отряда хищных.
Другой пример. Класс млекопитающих характеризуется определенным набором основных признаков и свойств, отличающих их от рептилий. Этот набор признаков (постоянная теплокровность, живорождение, вскармливание детенышей молоком, развитие мозга, особенности локомоции, строение органа слуха и другие особенности) возник, как показывают палеонтологические данные, не у одного вида или группы близких видов, а в разных стволах одного пучка филогенетически близких групп рептилий, причем не одновременно (см. рис. 17.2). Тем не менее весь класс млекопитающих надо считать монофилетической группой; он возник от одного таксона того же ранга (от класса рептилий).
Сейчас много примеров того, как по мере накопления данных по морфологии, палеонтологии, особенностям образа жизни отдельных групп их приходится разделять на самостоятельные. Так было с отрядом грызунов (Glires), в который еще недавно включали в качестве подотряда зайцеобразных. Теперь нет сомнения, что сходство зайцеобразных с грызунами внешнее, конвергентное, и ныне они выделены в самостоятельные отряды Lagomorpha и Rodentia, имеющие разное происхождение. Еще недавно в один отряд хищных птиц объединяли соколов и сов — две совершенно различные по происхождению группы птиц. Как только выясняется полифилетический характер таксонов, их следует разделять.
Однако проблема монофилии не исчерпывается признанием концепций широкой монофилии (происхождением одного таксона от другого такого же систематического ранга). В гл. 13 приводились некоторые из многочисленных примеров гибридогенного видообразования. Многие виды ириса, табака, полыни, костра, мятлика и других растений имеют гибридогенное происхождение (путем аллополиплоидии — гибридизации с последующим удвоением числа хромосом). Это означает, что на микроэволюционном уровне полифилия возможна. Возможность возникновения нового рода посредством гибридизации, т. е. полифилетически, доказана экспериментально (см. рис. 6.28). Сейчас известен ряд природных межродовых гибридогенных форм. Среди них рябинокизильник (Sorbocotaneaster), распространенный на юге Якутии, ячмень-клинэлимусовый гибрид (Hordeum x Clinelymus), распространенный на Памире, гибрид алычи и абрикоса — в Молдове.
Интересными примерами гибридогенного видообразования служат некоторые виды амфибий (из группы европейских зеленых лягушек) и рептилий (кавказские скальные ящерицы). В последнем случае видообразованию способствует возникновение партеногенетических форм.
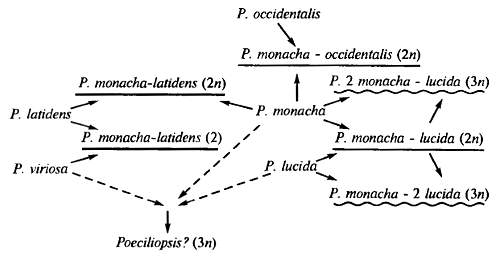
Рис. 20.3. Сетчатая эволюция в гибридогенном комплексе диплоидно-триплоидных, одно- и двуполых рыб рода Poecciliopsis. Однополые формы гибридного происхождения не подчеркнуты, диплоидные гибридогенные подчеркнуты, триплолидные гиногенетические подчеркнуты волнистой линией. В скобках указана степень полиплоидности однополых форм (по Б.В. Васильеву, 1985)
Итак, существующие генетические механизмы (аллополиплоидия, партеногенез) позволяют в определенных случаях прорвать барьер внешней стерильности, характерный для видовых форм в природе; это и ведет к полифилетическому возникновению видов. В этих случаях так называемое сетчатое родство (рис. 20.3), которым связаны все популяции внутри вида, обменивающиеся генетической информацией, оказывается частично характерным и для групп видового ранга. В последние десятилетия анализ происхождения каждой крупной филогенетической группы превращает ее из древа в куст. По-видимому, завоевание каждой крупной адаптивной зоны (основа возникновения последующего процесса аллогенеза — образования многих дочерних форм с частными адаптациями) было связанно с тем, что в гомологичных участках генома не одного, а разных видов подхватывались отбором гомологичные мутации. Поэтому «переход в другую адаптивную зону ... может затрагивать одновременно не один, а многие виды, принадлежащие к разным семействам и отрядам исходного класса» (Н.Н. Воронцов, 2001, с. 515, 517). Такой параллельный выход нескольких видов в новую зону и образование там новой крупной систематической группы называется парафилией. Парафилитическое происхождение предполагается не только для млекопитающих (см. рис. 17.2), но и для голосеменных, и покрытосеменных. Впрочем, как обращает внимание Н.Н. Воронцов, в основе парафилии все же лежит дивергенция: сама возможность гомологичного мутирования определяется общностью происхождения.
Другим фактором, заставляющим по-иному взглянуть на всеобщность и обязательность проявления принципа монофилии в возникновении природных таксонов, является существование огромной и разнообразной группы лишайников. Известно, что лишайники — своеобразные комплексные организмы, которые состоят из зеленых водорослей и грибов аско- или базидиомицетов. Образующие лишайники грибы не могут жить без водорослей. Существует около 10 000 видов лишайников, выделяемые как особый класс Lichenes. При этом возникает своеобразная ситуация: одновременно существуют филогенетические системы, включающие порознь водоросли, с одной стороны, а аско- и базидиомицеты — с другой. В то же время существует особая, самостоятельная филогенетическая система для лишайников. В целом как систематическая группа организмов они имеют полифилетическое происхождение.
При обсуждении проблемы полифилии приходится вспомнить и гипотезу о симбиогенном происхождении основных групп современных организмов путем последовательного объединения аэробных бактерий и микоплазм (см. гл. 5). Это позволяет говорить о полифилетическом происхождении типов и царств живой природы.
Итак, разнообразие эволюционных путей живой природы оказывается большим, чем это предполагалось в 50-е гг. XX века: в некоторых специальных случаях в процессе эволюции могли на разных уровнях вторично возникать полифилетические таксоны.
