20.2. Направленность и ограниченность эволюционного процесса
Естественный отбор может действовать по любому признаку или свойству, изменяя любой наследственный признак в чреде поколений. Однако было бы неверным на основании этого делать вывод о безграничности возможностей эволюции конкретной группы.
В проблеме направленности два основных аспекта: собственно проблема направленной эволюции и проблема реально существующих ограничений в эволюции того или иного филума. Рассмотрим эти направления более подробно.
В конце XIX в. Т. Эймер выдвинул концепцию ортогенеза (прямолинейное развитие), которую он противопоставил теории Ч. Дарвина. Согласно ей (впоследствии с близкой системой взглядов — концепцией номогенеза — выступил Л.С. Берг), эволюция осуществляется не на основе отбора случайно возникающих наследственных уклонений, как предполагал Ч. Дарвин, а на основе реализации неких внутренних законов (от греч. «nomos» — закон). Такая эволюция (номогенез) была противопоставлена дарвиновской эволюции как эволюции на основе случайности (тихогенез).
Сейчас можно сказать, что подобное противопоставление оказалось неточным уже потому, что статистическая природа многих явлений жизни (в том числе и эволюционных закономерностей) определяет важную роль случайности и не исключает, а предполагает проявление необходимости.
Говоря о развитии на основании определенных «внутренних законов», можно привести множество примеров, вроде бы подтверждающих прямолинейный характер эволюции: например, от пятипалого предка до однопалой современной лошади можно провести прямую линию, внешне как бы выражающую реализацию «внутренней закономерности» потери пальцев ноги (см. рис. 6.4).
Среди других примеров якобы направленности эволюции — редукция размеров и упрощение строения у однолетних трав, уменьшение размера тела островных животных по сравнению с их материковыми предками, усиление закрученности у некоторых моллюсков в позднем олигоцене и великое множество подобных эволюционных трендов, проявляющихся в ходе филогенеза разных групп независимо друг от друга.
Адаптации, внешне производящие впечатление направленности, реализуются в ходе эволюции всех достаточно подробно изученных филогенетических стволов (см. рис. 6.5).
Теория отбора дает объяснение всем этим случаям: на каждом из этапов эволюции естественный отбор оставлял особи с особенностями, лучшими в определенных условиях. Если условия существования группы сохранялись стабильными достаточно длительное время, направление действия отбора в ряду поколений также оставалось стабильным. Именно отбор направлял эволюцию по определенному руслу. Внешняя направленность, выражающаяся в развитии определенных особенностей строения в ходе эволюции, обусловлена не имманентными «внутренними» закономерностями, а действием отбора.
Однако у проблемы направленности в эволюции есть и второй аспект: существование тех или иных эволюционных запретов. Например, у червеобразного существа практически нет онтогенетических предпосылок для формирования крыльев, но такие предпосылки есть у млекопитающих (в виде подвижного скелета конечностей и подвижной кожи). У человека нет онтогенетических предпосылок для формирования кисточки на кончике хвоста, потому что у него нет развитого хвоста, но такие предпосылки есть у других млекопитающих с развитым хвостом. Изменчивость признаков и свойств у любого вида живых организмов всегда ограничена.
Выяснение системы запретов в эволюции той или иной группы является перспективным подходом предвидения возможных путей эволюции группы в дальнейшем. Часто такие системы запретов бывают связаны с некоторыми физико-химическими или структурно-механическими свойствами живых организмов. Поясним это аналогией со строением кристаллов в неорганической природе. Установлено, что в природе возможно существование только 231 формы кристаллических решеток (расположение структурных компонентов внутри кристалла). И все существующее разнообразие кристаллов ограничено числом вариантов, определяемых физико-химическими свойствами атомов и молекул. Аналогичные системы запретов существуют в живой природе. Необходимость снабжения листа зеленого растения питательными веществами, оттока синтезированных продуктов в другие части растения, а также необходимость механической поддержки листовой пластинки определяет систему структурно-механических запретов в эволюции формы листовой пластинки, включающую все возможные варианты формы листа как органа фотосинтеза (рис. 20.1).
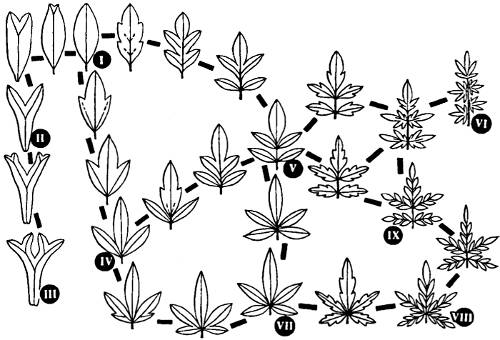
Рис. 20.1. Взаимоотношения всех возможных вариантов формы листовой пластинки. Пример ограничений в эволюции, связанных со структурно-механическими запретами. I—IX — последовательные варианты формы листьев (рис. С.В. Мейена)
Ярким выражением системы запретов является закон (точнее — правило) гомологических рядов в наследственной изменчивости (Н.И. Вавилов, 1922): «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов». Такие «сходные ряды наследственной изменчивости» являются своего рода эволюционными каналами, которые ограничивают и как бы канализируют эволюционный процесс. Невольно напрашивается аналогия с процессом канализации онтогенеза, рассмотренном в гл. 14 (см. рис. 14.5).
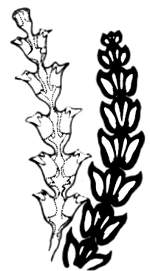
Рис. 20.2. Возможно, в конструкции скелетных элементов колонии гидроидных полипов Diphasia и шишки туи (Thuja) выражаются какие-то неизвестные нам пока общие системы структурно-механических запретов (из А. Портмана, 1961)
Выражением существования неких объективных эволюционных запретов является и то обстоятельство, что природа за миллиарды лет эволюции не «изобрела» колеса как эффективного способа передвижения. Это произошло потому, что существует определенная система структурно-механических запретов. Выяснение таких систем запретов — не простое дело, как может показаться на первый взгляд (рис. 20.2). Известно, например, что акуловые рыбы могут жить на значительных глубинах и в поверхностных слоях воды, в открытом море и на мелководье, но не могут жить в пресных водах — у них иной тип водно-солевого обмена со средой, нежели у костистых рыб. Налицо существование запрета, основанного на морфофизиологических особенностях организации группы. Однако известны два (!) вида акул, способных жить в пресных водах. Каким-то образом в процессе эволюции здесь система запрета оказалась преодоленной. Изучение таких исключений из правила чрезвычайно интересно. Они — свидетельство необычных путей эволюционного процесса.
Чем выше ранг таксона, тем меньше в целом число запретов. Млекопитающие как класс освоили все имеющиеся на планете адаптивные зоны, тогда как отдельные отряды млекопитающих в процессе специализации приобрели многочисленные системы запретов, препятствующие проникновению в ту или иную зону (например, китообразные — систему запретов жизни на суше, рукокрылые — жизни в почве и т.п.).
Итак, в основе направленности эволюции лежит, с одной стороны, направленное действие естественного отбора на протяжении значительного числа поколений, а с другой стороны, система запретов, обусловленная организацией данной группы. Однако, как многократно подчеркивалось ранее, любые морфофизиологические, онтогенетические и генотипические особенности сами по себе — результат предшествующей эволюции (и действия отбора в прошлом).
Таким образом, современная эволюционная теория не только не отвергает явление направленности в эволюционном развитии, но, напротив, подчеркивает его как необходимое следствие дарвиновского понимания характера эволюции.
