10.6. Эффективность и скорость действия естественного отбора
Эффективность и скорость действия естественного отбора зависят в первую очередь от величины давления отбора (т. е. степени количественного воздействия). Для оценки величины давления отбора необходимо познакомиться с понятием адаптивная ценность генотипа.
В процессе отбора индивидуальные достоинства каждого генотипа в популяциях оцениваются по способности передавать гены будущим поколениям. Способность генотипа к выживанию и воспроизведению по сравнению с другими генотипами в популяциях называется адаптивной ценностью генотипа. Она характеризует степень относительной приспособленности генотипа (W), которая колеблется в пределах от 0 до 1. При W = 0 передача генетической информации индивида следующему поколению отсутствует и аллель исчезает из популяции; при W — 1 образуется максимальное число гамет с данным наследственным признаком (полностью реализуются потенциальные возможности к размножению).
Адаптивная ценность генотипа определяется всем комплексом генов, поэтому даже генотипы, гомозиготные по одному и тому же гену, могут обладать различной адаптивной ценностью. Количественно адаптивную ценность выражают в % частот аллелей до и после отбора, т. е. с учетом их частот в поколениях, что, в свою очередь, зависит от коэффициента отбора.
Коэффициент отбора (S) характеризует интенсивность элиминации или снижение воспроизведения мутантного аллеля по сравнению с исходной формой. Представляет величину, обратную адаптивной ценности генотипа: варьирует от 1 до 0. Чем больше адаптивная ценность генотипа, тем ниже коэффициент отбора. Если W = 1, то S = 0. Если из 100 родившихся особей с определенным признаком до размножения доживут все, а с другим признаком — лишь 99 особей, то можно сказать, что в первом случае коэффициент отбора равен 0, а во втором — 0,01. В природных условиях коэффициент отбора обычно не превышает 0,10—0,20, чаще имеет меньшие значения. Конечно, по отношению к летальным мутациям, резко понижающим жизнеспособность организмов, коэффициент отбора может достигать 1,0.
Коэффициент отбора может выражать как возрастание, так и убывание концентрации аллеля. В первом случае имеет место размножение носителя, во втором — его элиминация.
В природе оценка адаптивной ценности генотипа по отдельным признакам происходит редко, обычно отбор идет по комплексу признаков. Для характеристики адаптивной ценности всех генотипов в популяции определяют среднюю приспособленность W. Она равна сумме частот генотипов после отбора и определяется по формуле Харди — Вайнберга (см. гл. 8):
W = p2W0 + 2pqW1 + q2W2.
При этом среднюю приспособленность доминантной гомозиготы принимают за 1, а приспособленность остальных генотипов соотносят к ней в процентах:
| Генотипы | Средняя приспособленность |
| AA | W0 = W0/W0= 1 |
| Aa | W1 = W1/W0 = 1—S |
| aa | W2= W2/W0 = 1— S2 |
Применяя эти формулы и зная соотношение генотипов до и после отбора, легко определить коэффициент отбора (табл. 10.3).
| Таблица 10.3. Пример определения величины коэффициента отбора (S) генотипов | |||
| Показатели | Генотипы | ||
| AA | Aa | aa | |
| Частота до отбора | 0,25 | 0,50 | 0,25 |
| Частота после отбора | 0,35 | 0,48 | 0,17 |
| Относительная выживаемость | W0 = 0,35/0,25 = 1,4 W0/W0 = 1,4/1,4 = 1,0 |
W1 = 0,48/0,50 =0,96 W1/W0 = 0,96/1,4=0,7 |
W2 = 0,17/0,25 = 0,68 W2/W0 = 0,68/1,4=0,4 |
| Коэффициент отбора | 1,0—1,0 = 0 | 1,0 —0,7 = 0,3 | 1,0—0,4 = 0,6 |
Чем значительнее коэффициент отбора, тем интенсивнее отбор против каких-либо генотипов, т. е. выше давление отбора. Давление отбора за отрезок времени всегда приводит к достижению определенного положительного результата (изменению частот генов). Это эффективность отбора.
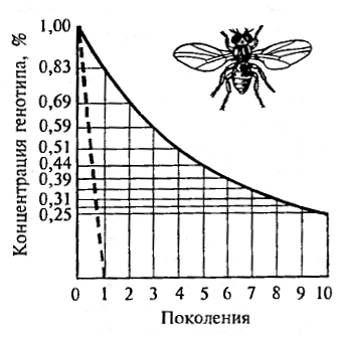
Рис. 10.6. Отбор против доминантного (пунктир) и рецессивного генотипа при давлении отбора S = 1. Исходная концентрация генотипов равна 1% (из Н.П. Дубинина, 1970)
Отбор особенно эффективен против доминантных мутаций и при условии их полного выражения (экспрессивность) и проявления (пенетрантность, см. 8.2). При 5=1 популяция за одно поколение избавляется от доминантных летальных мутаций (рис. 10.6). Например, отсутствие насыщения популяций человека мутацией одной из форм карликовости (ахондроплазия) объясняется ее доминантностью и устранением в результате отбора. Особи, несущие ее, умирают в раннем возрасте, и по этой причине она не накапливается в популяциях человека. Отбор против рецессивных генов и при неполном доминировании, наоборот, затруднен. В гетерозиготном состоянии (под «прикрытием» нормального фенотипа) они ускользают от действия отбора. Особенно трудно избавить от рецессивных мутаций большие популяции, где их перевод в гомозиготное состояние происходит очень медленно (рис. 10.7).
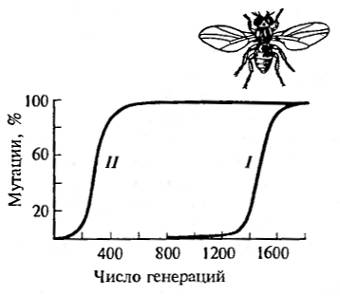
Рис. 10.7. Отбор против доминантных (II) и рецессивных (I) признаков при низких и высоких концентрациях (по В. Людвигу, 1940)
Эффективность отбора, как следует из данных рис. 10.7, в значительной степени зависит от исходной концентрации гена в популяции. При очень низких (I) и очень высоких (II) концентрациях отбираемого признака (гена) отбор действует слишком медленно, при средних концентрациях — очень быстро даже при низких значениях коэффициента отбора. Расчеты показывают, что при значении коэффициента отбора, равном всего 0,01, в популяции с доминантным геном А и рецессивным а возрастание частоты первого гена пойдет следующим образом (К. Пэтау, 1939):
| Возрастание, % | Число поколений |
| 0,1—1 | 232 |
| 1—50 | 559 |
| >50—98 | 5189 |
| 98—99 | 5070 |
| 99—99,9 | 90 231 |
Заметим, что данные расчеты значительно упрощены, в них не учтено давление других факторов на концентрацию гена в популяции (например, мутационного процесса, отбора по другим коррелированным с изучаемым признаком и т. п.).
Отсутствие отбора (5 = 0) вряд ли возможно в природных популяциях. Различные генотипы обязательно отличаются друг от друга, и при постоянном давлении жизни у них разные вероятности достижения репродуктивного возраста.
