2.7. Царство растений
Хотя жизнь на нашей планете, вероятно, зародилась примерно 3,5 млрд. лет назад, первые организмы заселили сушу не ранее, чем 420 млн. лет назад. Это были самые первые растения. Растения – автотрофные эукариоты, которые адаптировались к жизни в воздушной среде. Единственные другие автотрофы среди эукариот – это водоросли, специализировавшиеся к жизни в воде. Напоминаем: автотрофами называют организмы, использующие в качестве источника для синтеза органических веществ неорганический углерод, а именно диоксид углерода. Однако помимо источника углерода для такого синтеза требуется еще и энергия (разд. 2.3.4), и растения, будучи фотоавтотрофами, используют в качестве источника энергии свет. Этот способ питания более принято называть фотосинтезом.
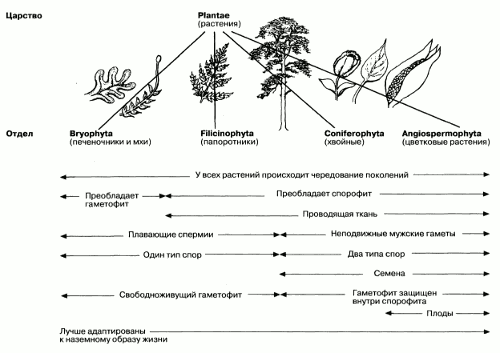
Рис. 2.33. Систематика растений и некоторые основные тенденции в эволюции растений.
История эволюции растений – это постепенно совершенствующаяся адаптация к жизни на суше. Именно эта история и составит одну из основных тем в нашем изучении растений. Систематика растений, рассматриваемых в данной книге, показана на рис. 2.33. Там же приводится краткий перечень некоторых основных тенденций в эволюции растений, связанных с адаптацией к жизни на суше, которые также будут рассматриваться в этом разделе.
2.7.1. Отдел Bryophyta (печеночники и мхи)
Печеночники и мхи – наиболее примитивные из всех наземных растений. Полагают, что они произошли от зеленых водорослей. В отдел Bryophyta (мохообразные) входят два главных класса – Hepaticae (печеночники) и Musci (мхи). Обе эти группы плохо приспособлены к жизни на суше, и поэтому они привязаны к сырым затененным местам. Систематика и основные признаки мохообразных приведены в табл. 2.8.
Таблица 2.8. Систематика и особенности отдела Bryophyta (моховидные)
| Отдел Bryophyta | |
| Общие признаки Чередование поколений, при котором преобладает гаметофитное поколение Нет проводящей ткани, т.е. нет ни ксилемы, ни флоэмы Тело представлено талломом или слегка дифференцировано на простые «листья» и «стебли» Нет настоящих корней, стеблей и листьев; гаметофит прикрепляется к субстрату нитевидными ризоидами Спорофит прикреплен к гаметофиту, полностью зависит от него и питается за его счет Споры образуются на спорофите в споровой коробочке, расположенной на конце тонкой ножки, возвышающейся над гаметофитом Встречаются главным образом в сырых затененных местах |
|
| Класс Hepaticae (печеночники) |
Класс Musci (мхи) |
| Гаметофит представлен упрощенным образованием, форма которого варьирует от слоевищной (редко) до «облиственной» со стеблем (у большинства видов); имеются переходные формы дольчатого типа «Листья» (у облиственных печеночников) расположены вдоль стебля тремя рядами Ризоиды одноклеточные Для рассеивания спор коробочка спорофита раскрывается на четыре створки; рассеиванию спор помогают элатеры ПРИМЕРЫ: Pellia – слоевищный печеночник, Marchantia – слоевищный печеночник; антеридии и архегонии расположены на своеобразных стебельках, возвышающихся над талломом Lophocolea – облиственный печеночник; обычно встречается на гниющей древесине |
Гаметофит «облиственный», имеет «стебель» «Листья» расположены спирально Ризоиды многоклеточные Рассеивание спор из коробочки спорофита происходит с помощью сложного механизма, срабатывающего в сухую погоду и включающего участие зубцов и пор ПРИМЕРЫ: Funaria Mnium – обыкновенный лесной мох, похожий по внешнему виду на Funaria Sphagnum – болотный мох; во влажной кислой среде (на болотах) образует слои торфа |
Bryophyta – небольшие растения с несложным строением. Опорная и проводящая ткани у них развиты слабо или же вовсе отсутствуют. Нет у них дифференцированной ксилемы и флоэмы, так же как нет и настоящих корней. В почве они удерживаются тончайшими нитями, называемыми ризоидами. Вода и минеральные соли поглощаются всей поверхностью тела, в том числе и ризоидами. А это означает, что в отличие от настоящих корней ризоиды служат лишь для закрепления растения в грунте. (В настоящих корнях, как и в настоящих стеблях или листьях имеются проводящие ткани.) Мохообразные не покрыты сверху кутикулой, либо кутикула столь тонка, что она не препятствует потере (или поступлению) воды. Тем не менее многие мохообразные приспособились выдерживать засушливые периоды, используя для этой цели какие-то не совсем понятные механизмы. Например, было установлено, что такой хорошо известный ксерофитный мох, как Grimmia pulvinata, больше года остается живым при 20° C в абсолютно высушенном состоянии. Сразу же после того, как растение попадает во влажную среду, у него восстанавливаются все функции.
Чередование поколений
Как у всех наземных растений и некоторых наиболее высоко организованных водорослей, таких как ламинария, у мохообразных наблюдается чередование поколений. В ходе жизненного цикла происходит смена двух типов организмов: поколение гаплоидного гаметофита и поколение диплоидного спорофита поочередно сменяют друг друга, что схематически показано на рис. 2.34. Гаплидное поколение называется гаметофитом (от греч. gamete – жена, gametes – муж; phylon – растение), так как оно способно к половому размножению и образует гаметы. Поскольку образование гамет происходит в результате митоза, они тоже гаплоидны. Сливаясь, гаметы образуют диплоидную зиготу, из которой вырастает следующее поколение – поколение диплоидных спорофитов. Они называются спорофитами, потому что способны к бесполому размножению с образованием спор. Споры образуются в результате мейоза, т.е. здесь происходит возврат к гаплоидному состоянию. Гаплоидные споры дают начало гаметофитному поколению. Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла; это поколение называют доминантным. У моховидных доминирует поколение гаметофитов, у всех остальных наземных растений – поколение спорофитов. Доминирующее поколение принято помещать в верхнюю половину схемы, изображающей жизненный цикл.

Рис. 2.34. Обобщенная схема жизненного цикла растений, отражающая чередование поколений. Обратите внимание на наличие гаплоидных (n) и диплоидных (2n) стадий. Гаметофит всегда гаплоидный и всегда образует гаметы путем митотического деления. Спорофит всегда диплоидный и всегда образует споры в результате мейотического деления.
Внимательно изучите рис. 2.34, так как на нем в обобщенном виде представлен жизненный цикл всех наземных растений, в том числе и наиболее высокоорганизованных цветковых растений. Никогда не забывайте, что гаметы у растений образуются не в результате мейоза, как у животных, а в результате митоза; мейотическое деление происходит при образовании спор.
Класс Hepaticae – печеночники
Характерные признаки класса Hepaticae представлены в табл. 2.8. По своему строению печеночники намного проще, чем мхи, и в целом они более привязаны к сырым и затененным местам. Их можно найти на берегу рек и ручьев, на влажных камнях и среди болотной растительности. У большинства печеночников видны правильные доли или вполне выраженные «стебли» с небольшими простыми «листочками». Проще всего устроены талломные печеночники, тело которых представлено плоским слоевищем и не разделено на стебли и листья.
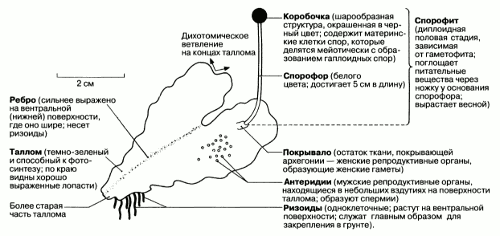
Рис. 2.35. Внешние признаки Pellia (печеночника). Гаметофит изображен вместе с прикрепленным к нему несамостоятельным спорофитом.
Примером может служить Pellia, печеночник, широко распространенный по всей Великобритании. Это растение окрашено в тускло-зеленый цвет, ширина плоских «веточек» составляет около 1 см. Внешние признаки Pellia приведены на рис. 2.35.
Класс Musci – листостебельные мхи
Основные признаки листостебельных мхов перечислены в табл. 2.8. Листостебельные мхи гораздо более дифференцированы, чем печеночники, но, как и печеночники, это небольшие растения, встречающиеся главным образом в сырых местах. Они часто образуют плотные подушки.
Funaria – обычный представитель лиственных мхов на полях, вырубках и перекопанных землях, где она поселяется одной из первых. Funaria особенно любит селиться на кострищах и пожарищах. Это один из самых обычных сорняков в теплицах и садах. Внешний вид Funaria показан на рис. 2.36.
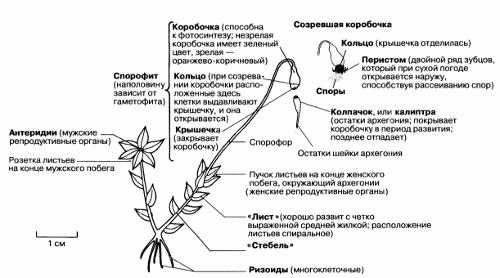
Рис. 2.36. Внешние признаки мха Funaria. Гаметофит изображен вместе с прикрепленным к нему наполовину самостоятельным спорофитом.
Как и у печеночников, для оплодотворения Funaria необходима вода. Когда поверхность таллома увлажняется, созревшие антеридии поглощают воду и лопаются, высвобождая на поверхность мужские гаметы (спермии). Каждый спермий снабжен двумя жгутиками. Спермии подплывают к архегониям, в каждом из которых находится женская гамета, или яйцеклетка. Оплодотворение, т.е. слияние ядра спермия с ядром яйцеклетки, происходит в архегонии. В результате слияния образуется диплоидная зигота, которая, вырастая из архегония, дает начало новому спорофиту.
