Переходные формы. Несмотря на то, что между крупными естественными группами животных, растений и микроорганизмов, как правило, существуют глубокие разрывы, вызванные вымиранием промежуточных форм, в ряде случаев мы обнаруживаем переходные формы. Существование форм, сочетающих в своем строении признаки разных типов организации и занимающих поэтому промежуточное систематическое положение, определяется общим родством организмов. При таком родстве между отдельными, далеко отошедшими друг от друга крупными ветвями древа жизни могут существовать мелкие ветви, носящие промежуточный характер (рис. 6.23).
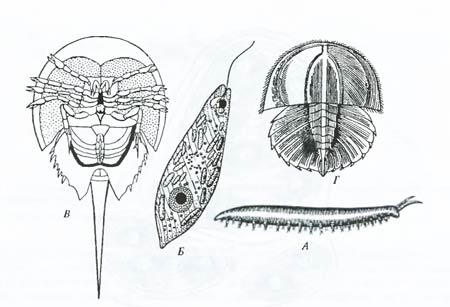
Рис. 6.23. Примеры ныне существующих форм: А – перипатус (Peripatus laurocerasus), несущий признаки членистоногих и кольчатых червей; Б – эвглена (Euglena viridis), соединяющая признаки животных и растений; В – мечехвост (Limulus polyphemus), занимающий промежуточное положение между современными типичными членистоногими и ископаемыми трилобитами; Г – личинка мечехвоста, похожая на личинку трилобита (по И.И. Шмальгаузену, 1969)
Одним из примеров переходных форм между оболочниками (наиболее примитивной группой хордовых) и позвоночными животными служит род ланцетников, для которых характерны все основные признаки хордовых, но развиты они незначительно (Л.О. Ковалевский).
Существование промежуточных форм в современном органическом мире – свидетельство единства организации крупных стволов древа жизни и единства их происхождения.
Микросистематика. Начиная с классических работ П.П. Семенова-Тян-Шанского в случае хорошо изученных групп видов возникала возможность выявления их внутривидовой структуры – подвидов, рас и других группировок (микросистематика). Такое изучение имеет важное значение для понимания особенностей микроэволюции. Сейчас ясно, что в системе вида могут существовать сложные иерархические взаимоотношения популяций и их групп.
При этом выявление истинного филогенетического родства таких группировок, необходимое для определения внутривидовых таксономических категорий, оказывается в то же самое время и восстановлением их микрофилогенеза – путей исторического развития отдельных частей видового населения и вида в целом (рис. 6.24).
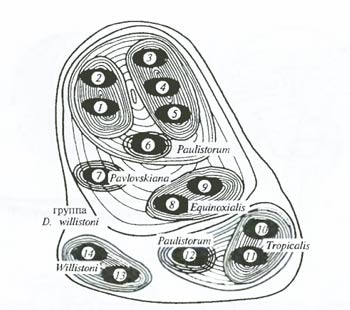
Рис. 6.24. Микрофилогенез дрозофил группы Willistoni можно восстановить на основании экспериментов по скрещиванию, анализа строения хромосом и биохимического полиморфизма. Иерархия филогенетических форм в данном случае должна быть описана так: популяция – подвид – полувид – вид – надвид (по Б. Спасскому и др., 1971)
Drosophila willistoni представлена двумя географически изолированными подвидами (13, 14), вид D. equinoxialis также представлен двумя подвидами (8, 9) с еще более глубокой репродуктивной изоляцией между ними: вид D. paulistonim состоит из шести полувидов (группы популяций 1 и 2, 3 и 5 обитают симпатрически, между ними существует полная репродуктивная изоляция в природе, но особи этих групп из разных местностей могут скрещиваться в лаборатории). Представители всех шести видов не скрещиваются ни в природе, ни в лаборатории (редкие лабораторные гибриды всегда стерильны). В то же время различия между D. willistoni и остальными пятью видами больше, чем между остальными пятью видами
Популяционная морфология. А. Уоллес в XIX в. приводил вариационные ряды значений исследуемых признаков для небольших групп особей. С распространением популяционного мышления (см. гл. 7) морфологические исследования также оказались удобным оружием для изучения текущих процессов микроэволюции. Популяционно-морфологические методы, позволяют улавливать направления естественного отбора по изменению характера распределения значений признака в популяции на разных стадиях ее существования или при сравнении разных популяций (рис. 6.25, А, Б).
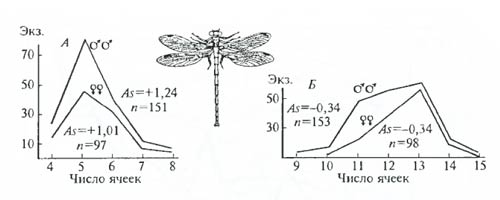
Рис. 6.25. Асимметрия в распределении значений количественного признака может показать направление давления естественного отбора (по А.В. Яблокову, 1966).
Число ячеек на определенных участках крыла стрекозы Lestes sponsa
Морфологические методы позволяют выделять среди однородной группы признаков те, которые находятся под большим или меньшим давлением со стороны отбора (рис. 6.26), хотя в этом случае само адаптивное значение признака может быть вскрыто лишь дополнительными эколого-физиологическими исследованиями. Одной из трудностей в применении морфологических методов к изучению процесса микроэволюции является сложность разграничения действия современных эволюционных факторов от изменчивости, определяемой широкой нормой реакций. Так, получивший одно время широкое распространение в нашей стране метод морфофизиологических индикаторов (С.С. Шварц), позволяющий различать физиологическое состояние популяций животных и направления ее адаптивной перестройки, часто оказывался скомпрометированным тем обстоятельством, что за эволюционно значимые индикаторы принимались мимолетные, кратковременные и обратимые изменения.
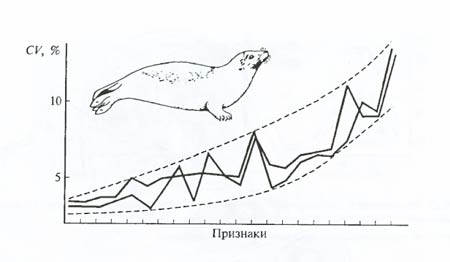
Рис. 6.26. Значения коэффициента вариации, резко выпадающие из «потока изменчивости» группы признаков, могут показывать на отношение данного признака к давлению отбора. В черепных промерах гренландского тюленя (Pagophilus groenlandica) пониженным значением коэффициента вариации отличаются размеры слуховой кости, а повышенной – носовой кости. Первый признак крайне важен и жестко контролируется отбором во все периоды онтогенеза, второй – малозначим (по А.В. Яблокову, 1966)
Изучение микроэволюционного процесса с помощью морфологических методов является содержанием нового направления в современной морфологии – популяционной морфологии, дисциплины, тесно связанной с популяционной генетикой и экологией.
