Генетики первой половины XX в. о механизмах надвидовой эволюции
Мутационная теория эволюции Гуго де Фриза (Н. de Vries), созданная в 1901-1903 гг., широко известна. Исследуя длительное время изменчивость в роде энотера и у других растений, де Фриз обнаружил явление спорадического возникновения среди линнеевских видов особей с необычным новым признаком, стойко передающимся потомству в неограниченном числе поколений. Для обозначения этого явления он предложил термин «мутация», ставший азбучным понятием в генетике. Излишне пояснять, что генетический термин «мутация» не имеет ничего общего с мутацией Ваагена – понятием, ранее введенным в палеонтологию.
Зарегистрировав появление константных мутантных форм, де Фриз получил основание рассматривать обычные линнеевские виды, или виды систематиков, как смеси монотипичных, генетически далее неразложимых элементарных видов (жорданонов), отличающихся друг от друга одним наследственным признаком. На превращениях именно таких элементарных видов де Фриз и построил свою мутационную теорию эволюции.
По этой теории эволюционные преобразования совершаются скачкообразно. Время от времени под действием пробудившейся «созидательной силы» природы тот или иной вид, дотоле длительное время пребывавший в неизменном состоянии, испытывает мутацию и практически внезапно превращается в новый вид, который теперь сразу и на столь же долгое время, как и его предшественник, становится постоянным. Периоды покоя, длящиеся тысячелетиями, чередуются с мутационными периодами, кажущимися по сравнению с первыми всего лишь мгновениями.
Де Фриз отмечает, что мутации, а следовательно, и образование новых видов «не зависят от жизненных условий» (Фриз де, 1904. С. 199). Видообразование совершается также независимо от естественного отбора. «Для этого, – по его словам, – не нужно ни ряда поколений, ни борьбы за существование, ни удаления негодных особей, ни подбора» (там же, с. 196).
Борьба за существование в природе имеет место, но она идет не между особями одного вида, как полагал Дарвин, а между разными видами. При этом борьба за существование, в отличие от точки зрения Дарвина, не увеличивает видовое разнообразие, а сокращает его за счет выбраковки нежизнеспособных форм. Таким образом, Г. де Фриз свел сложный процесс видообразования и эволюции к одному фактору – «видообразовательной изменчивости» (Завадский, 1973. С. 274) – и тем самым избавил своих оппонентов от необходимости пространной критики своей концепции.
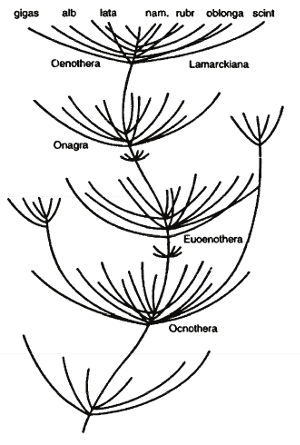
Рис. 3. Схема видообразования (по: де Фриз. 1904)
Де Фриз наглядно отобразил свою концепцию взрывного видообразования в виде схематичного родословного древа (рис. 3), внешне очень напоминающего хорошо знакомую аквариумистам водоросль кабомбу. Это длинная, местами ветвящаяся нить, образующая массу мутовок, находящихся на одинаковых расстояниях друг от друга. Каждая ветвь мутовки представляет собой вид или подвид, имеющий начало и конец, а каждое звено нити между соседними мутовками – неизменное состояние родоначального вида. Каждая мутовка соответствует мутационному периоду, все же древо в целом отображает род.
Этим де Фриз не ограничивается. Он добавляет, что рисунок можно было бы подобным же образом продолжить вниз – «вплоть до самых древних живых существ», и далее пишет: «На рисунке… мы восходим от видов к сборным видам (Oenothera lamarckiana), от сборных видов – к подродам (Onagra и Euoenothera), а отсюда – к родам (Oenothera). Более древним взрывам соответствовали бы подсемейства, семейства и все высшие системы. Если бы вся система была известна нам без пробелов и генеалогическое древо имело бы форму обыкновенной дихотомической таблицы для определения, то каждая точка разветвления означала бы для нас место взрыва…» (Фриз де, 1904. С. 192).
Сопоставив схему видообразования де Фриза с диаграммой дивергенции Дарвина, мы увидим, что они, равно как и трактовка обоими учеными способов осуществления надвидовой эволюции, принципиально сходны. В обоих случаях новые формы образуются путем ответвления от прежде единой родоначальницы. Правда, по диаграмме Дарвина, ветвление дихотомично, так как промежуточные формы вымирают; у де Фриза оно веерообразно (мутовчато), поскольку промежуточные формы, как и исходная, при отсутствии внутривидовой борьбы сохраняются. Но эти различия не столь принципиальны. Гораздо важнее иное различие. У Дарвина нарождение новых видов – плавный процесс, совершающийся медленно и постепенно, у де Фриза – резкий и скачкообразный.
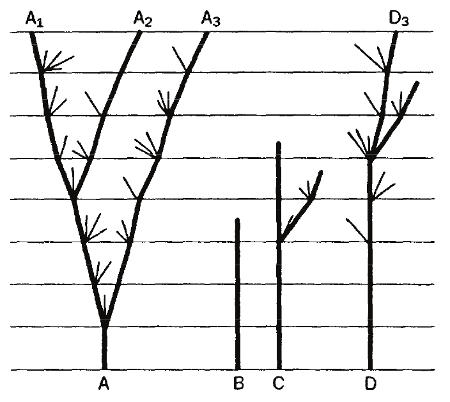
Рис. 4. Фрагмент диаграммы по Дарвину (только А) (из: Сковрон, 1987)
Однако в понимании способов надвидовой эволюции де Фриз и Дарвин едины. Для обоих образование высших таксонов – результат постепенного накопления изменений и, стало быть, дело времени. По де Фризу, каждый крупный эволюционный шаг складывается из серии мутаций. Никакого качественного различия между процессами видообразования и крупномасштабной эволюцией де Фриз, как и Дарвин, не проводит.
Подходя к оценке прогрессивного развития, связанного с «усовершенствованием организации», крайне упрощенно, де Фриз считал, что для осуществления эволюции от начала жизни до ее современных высших форм требовалось возникновение всего нескольких тысяч наследственных единиц, или эволюционных шагов. «Обыкновенно в каждый мутационный период, – писал де Фриз, – организация подвигается на один шаг. Следовательно, сколько таких шагов сделала организация с самого начала, столько было и мутационных периодов» (там же, с. 201–202).
Приняв продолжительность жизни на Земле равной 24 млн лет, а число мутационных периодов равным 6000, де Фриз нашел, что средняя продолжительность фазы покоя между мутационными периодами должна составлять 4000 лет (по замечанию самого де Фриза, это «очень грубое приближение»). Приведенные цифры характеризуют наиболее быстрый прогресс, который осуществили высшие растения и животные. Что касается низших, то у них число мутационных периодов было невелико, а фазы покоя весьма продолжительны. Де Фриз добавляет при этом, что если вслед за Дарвином объяснять прогрессивную эволюцию отбором и накоплением мелких изменений, то «не хватит… и миллиардов веков» (Фриз де, 1932. С. 70).
В первой половине геологической истории развитие мира живых существ должно было идти в несколько раз быстрее, чем в более позднее время. Мало-помалу прогресс ослабевал. «С появлением человека цель, кажется, была достигнута, и теперь все идет так лениво, что прогресс как будто закончен: нам кажется, что мы совершаем лишь вместе с ним его последние шаги» (Фриз де, 1904. С. 209). Этим соображением чисто финалистического характера де Фриз предвосхитил популярную среди финалистов 30-40-х годов концепцию цикличности и затухания эволюции.
Сколь неочевиден общий прогресс живого, связанный с «постепенным умножением числа свойств и признаков», прогрессивные мутации возникают, по мнению де Фриза, очень редко. В природе гораздо чаще встречаются мутации регрессивные («ретрогрессивные»), связанные, например, с выпадением признака, и «…происхождение видов в природе идет по большей части ретрогрессивным путем» (Фриз де, 1932. С. 124).
В теории де Фриза четко представлены два пути видообразования: 1) «групповой способ видообразования», при котором новые формы появляются сбоку главного ствола в виде веточек, образующих мутовку, и 2) «филогенетический», осуществляющийся на линиях, связывающих мутовки. По мнению де Фриза, первый способ видообразования обеспечивает богатство и разнообразие природы, а второй – «свойственное системе расчленение», но они не отделены друг от друга резкой гранью (Vries de, 1918; Фриз де, 1932. С. 118–119). В указанных способах видообразования легко угадывается близость к современным представлениям о кладистическом и филетическом видообразовании.
Завершая анализ теории де Фриза, необходимо сказать, что, несмотря на отдельные высказывания в поддержку эволюционной роли естественного отбора (Vries de, 1918), де Фриз явился автором в целом антидарвиновской, достаточно механистической и упрощенческой концепции, которую К.М. Завадский (1973) отнес к одной из основных разновидностей генетического антидарвинизма. В то же время нельзя не отметить, что, будучи беспристрастным и блестящим исследователем, наделенным большой научной интуицией, де Фриз высказал три кардинальных положения, составивших ядро современной теории прерывистого равновесия и находившихся еще недавно в фокусе острых дискуссий. К ним относятся идея периодичности мутирования и эксплозивности видообразования, отрицание внутривидовой борьбы и признание макроэволюционной роли межвидовой борьбы, положение об образовании большинства новых видов за счет боковых ответвлений.
Совершенно иного взгляда на соотношение внутривидовой и надвидовой эволюции придерживался другой генетик – современник де Фриза Л. Кено.
Л. Кено вошел в историю эволюционизма как автор теории преадаптации. Основная идея этой теории была высказана Кено уже в 1901 г. (Cuenot, 1901), т.е. до опубликования де Фризом мутационной теории, а затем оформлена в виде законченной доктрины в ряде публикаций (Cuenot, 1925, 1929, 1936). Кено с самого начала считал, что между условиями среды, в которой рождается новый вид, и его адаптацией к ней никакой причинной связи не существует и что особенности видовой организации возникают отнюдь не под действием ламарковских факторов или естественного отбора. Он с готовностью принял мутационизм, полагая, что эта концепция и его собственная теория дополняют друг друга, ибо путем мутаций, по его мнению, способны формироваться новые структуры. Не без гордости Кено причислял себя к самой молодой школе эволюционистов, которую назвал «менделистской, или мутационистской, или еще преадаптационистской» (Cuenot, 1921. Р. 467).
Со временем, однако, Кено становилось все более ясным, что при всем значении мутационного процесса как материальной базы эволюции его возможности ограничены рамками видообразования. Постепенно он приходит к выводу, что мутационная теория, дополненная фактором изоляции, способна удовлетворительно объяснить возникновение жорданонов (элементарных видов), географических рас и разновидностей, со временем достигающих видового уровня, но она ничего не может дать для понимания истинной (т.е. макро-) эволюции, характеризующейся появлением новых органов и морфологических структур. Отвергнув созидательную роль отбора, Кено настойчиво искал для объяснения этой эволюции крупного масштаба иные причины. В их поиске он все более склонялся к финализму (подробнее см.: Назаров, 1984).
Решающее значение в переходе Кено на позиции финализма имело, по-видимому, экспериментальное исследование возникновения в эмбриогенезе мозолистых затвердений на запястье передних конечностей африканской свиньи-бородавочника (Phacochoerus africanus), выполненное совместно с Р. Антони (Anthony, Cuenot, 1939). Именно в результате этой работы Кено уверовал в чудодейственный системный эффект одной крупной мутации, которая одновременно с созданием нового органа должна была породить и соответствующий инстинкт добычи пищи.
С этого момента Кено овладевает финалистическая идея о том, что сложные органы, подобные глазу позвоночных животных, крылу птицы или электрическим органам рыб, образуются не путем постепенных мелких и случайных изменений, аккумулируемых отбором, а только сразу, в результате одного неделимого акта под действием имманентной живому «зародышевой изобретательности». При этом новые органы должны с самого момента своего возникновения обладать полным совершенством. Прибегать к дарвиновскому способу объяснения с помощью мелких усовершенствований, по мнению Кено, в данном случае бессмысленно, поскольку до полного сформирования новые органы не способны функционировать и совершенно бесполезны.
К концу 40-х годов Кено окончательно разочаровывается в мутационной теории, ограничив сферу ее приложения расо- и видообразованием (микроэволюция). Отказывается он также и от теории преадаптации. Перейдя бесповоротно на позиции финализма, Кено теперь твердо считает, что крупномасштабная эволюция, связанная со становлением типов организации, управляется неизвестным нематериальным агентом психической природы и не имеет ничего общего с эволюцией в пределах вида.
Аналогичную метаморфозу во взглядах испытал также генетик и энергичный критик неоламаркизма и дарвинизма Э. Гийено. Из сказанного видно, что по вопросу о движущих силах внутривидовой и надвидовой эволюции единства не было уже среди самых зачинателей генетики. Его и трудно было бы ожидать, поскольку генетике как экспериментальной науке с ее длительное время остававшимся практически единственным методом гибридологического анализа оказалось недоступным изучение поведения признаков надвидовых таксонов, которые либо не скрещиваются, либо не менделируют. Исследователи периода развития классической генетики занимались преимущественно изучением распределения в потомстве менделистических признаков, характеризующих внутривидовые подразделения, в лучшем случае виды. Поэтому неудивительно, что их соображения о факторах надвидовой эволюции (если они вообще ею интересовались) были основаны в большей мере на умозрении, научной интуиции, чем на фактических данных. Однако и в этих условиях были высказаны догадки, намного опередившие уровень науки того времени.
