Основные и для всех очевидные различия между микро- и макроэволюцией усматриваются ими прежде всего в результатах: масштабы достигаемых морфофизиологических преобразований, а соответственно, и возникших при этом таксономических уровней резко различны. Несовпадение результатов непосредственно связано с временными характеристиками. Если процессы микроэволюции можно наблюдать на протяжении человеческой жизни, то процессы макроэволюции измеряются геологической шкалой времени. Микроэволюция может быть обратимой, макроэволюция необратима и уже в силу этого обладает определенной направленностью, а следовательно, и особыми закономерностями, проявляющимися в только ей свойственных феноменах (усложнение организации, смена этапов в развитии филогенетических групп, установление симбиотических отношений и т.д.). Некоторые исследователи указывают еще на одно различие, на которое обычно не обращается внимания и которое относится не только к области закономерностей, но и к самим механизмам эволюции: внутривидовая эволюция протекает в открытой генетической системе, надвидовая – в отдельных, изолированных друг от друга генофондах (Паавер, 1980). Следствием последнего обстоятельства оказывается возникновение иной формы отбора – межвидового, нуждающегося в особом анализе.
Большинство синтетистов справедливо признают, что в приложении теории естественного отбора к микроэволюции остается достаточное число спорных вопросов. К ним относится, например, трактовка отбора как дифференциальной смертности или дифференциальной размножаемости генотипов, способов оценки интенсивности отбора, соотношения ведущей и стабилизирующей форм отбора в эволюции и др. Недостаточно разработан вопрос о структуре самого естественного отбора. Если столь важные вопросы теории отбора как главного интегрирующего фактора эволюции остаются неясными на микроэволюционном уровне, то неудовлетворительное положение с оценкой его функциональной роли на уровне макроэволюции еще более усугубляется. Это можно подтвердить красноречивым признанием такого авторитетного теоретика синтетической теории, как К.М. Завадский. Сравнивая положение в учении о микроэволюции с положением в учении об арогенезе как важнейшем разделе теории макроэволюции, он отмечал, что «в теории естественного отбора существуют и неясности совсем другого рода. Все они связаны с применением этой теории к объяснению органообразования, возникновения новых типов организации и захвата новых зон жизни, т.е. к процессам арогенеза» (Завадский, 1972. С. 140). Правда, Завадский был убежден, что особенности движущих сил арогенеза состоят исключительно в специфичной интенсивности и длительности действия каждого из факторов микроэволюции, а также в их различном сочетании друг с другом.
Лет тридцать назад среди синтетистов выделилось крыло реформаторов (Уайт, Левонтин, Карсон, Буш, Гулд, Элдридж), которые предприняли попытку обосновать специфичность макроэволюции по такому важнейшему фактору, как наследственная изменчивость. В отличие от обычных генных и небольших хромосомных мутаций, служащих материалом для отбора в традиционной генетико-популяционной модели микроэволюции, они постулировали существование макромутаций как главного источника макроэволюционных новшеств, способных возникать сальтационно, вне связи с аккумуляцией мелких мутаций в случае микроэволюции. В СССР с подобной трактовкой специфичности макроэволюции выступил академик А.Л. Тахтаджян. Фактически на новом уровне знаний был совершен возврат к идеям Гольдшмидта. Вероятно, эволюционную платформу упомянутых исследователей следует рассматривать как крайний вариант синтетической теории, еще не претендующий на полную самостоятельность и не порывающий с ней генетической связи.
Наконец, надо сказать, что именно в связи с изучением проблем макроэволюции было высказано чрезвычайно важное соображение о канализирующем влиянии на ход макроэволюционных процессов экосистемных связей и отношений (Камшилов, Шварц, Чернов, Красилов, Одум и др.). Сама возможность появления и выживания новых видов есть, по-видимому, результат изменения биоценотической обстановки, а в конечном счете – сдвигов в режиме функционирования биосферы в целом (действующих через обратные связи). Эти соображения хорошо согласуются с представлением Дарвина о ведущей роли в эволюции биотических факторов. Поэтому вполне резонно считать, что причинные цепочки эволюции направлены не столько «снизу вверх» (от мутаций к популяциям и видам), сколько «сверху вниз» (от биоценоза к видам и популяциям). В учении о микроэволюции с подобным пониманием мы встречаемся редко.
При обсуждении вопроса о единстве факторов и движущих сил микро- и макроэволюции сразу возникает вопрос о доказательствах. В связи с невозможностью непосредственного наблюдения макроэволюции, ее экспериментального воспроизведения и моделирования этот вопрос приобретает особую остроту. Какого же рода обоснование данного тезиса предлагается в работах синтетистов?
Можно указать по крайней мере на четыре категории доказательств: выводы, полученные гипотетико-дедуктивным методом; экспериментальные данные об эффективности естественного отбора в гетерогенной (полиморфной) популяции; факты совпадения наследственных признаков близких видов с модификациями внутривидовых группировок; данные об арогенных популяциях. Характер доказательств первой категории ясен без особых комментариев. Это обычный метод построения научной теории, мало изменившийся со времен Дарвина. Экспериментальные данные об эффективности естественного отбора выполняют верификационную функцию. В ряде руководств по эволюционной теории (дарвинизму) приводятся одни и те же примеры. Таковы классические примеры «индустриального меланизма» у березовой пяденицы (Biston betularia) в странах Западной Европы, появление четырех рас ужей (Natrix sipedon) на островах озера Эри по рисунку поперечных полос, возникновение сезонных форм у погремка (Rhinantus major), превращение ярового рыжика (Camelina glabrata) в льняного (С. linicola) и др. В первых трех примерах (как и в большинстве других, здесь не приводимых) прослежены лишь начальные этапы микроэволюции, но отнюдь не видообразование, в случае которого можно было бы говорить о выходе на уровень макроэволюции. Относительно примера с рыжиком надо отметить, что постоянное возникновение модификационной формы (льняного рыжика) из ярового не означает, что наследственная форма (льняного) произошла тем же путем.
Таким образом, данные об эффективности естественного отбора, не достигшего уровни видообразовании, вовсе не доказывают, что микроэволюция приводит к макроэволюции. Что касается примеров внезапного видообразования, то они связаны в основном с хромосомными перестройками, полиплоидией и другими механизмами и свидетельствуют об эволюционных путях, минующих микроэволюцию.
Наконец, идея об арогенных популяциях, разрабатывавшаяся особенно интенсивно К.М. Завадским (1958, 1968, 1971), в теоретическом плане представляется нам достаточно плодотворной. Поскольку макроэволюционные события любого масштаба осуществляются только через видообразование, биологическая неравноценность возникающих видов или внутривидовых подразделений в перспективном отношении – очевидный факт. Однако само распознавание в природе арогенных или неарогенных популяций практически малоосуществимо. Получается, таким образом, что постулат о единстве микро- и макроэволюции прямых доказательств не имеет и опирается исключительно на доводы косвенного характера.
В 70-е годы XX в. среди советских эволюционистов распространился тезис, будто единственно возможный путь к познанию арогенной эволюции, а следовательно, и макроэволюции лежит через экспериментальное изучение механизмов микроэволюции и будто все другие подходы ненаучны. Тем самым заранее отвергалась целесообразность поиска возможной специфики в причинах и механизмах макроэволюции, игнорировались уже имевшиеся данные об иных путях надвидовой эволюции в некоторых группах растений и животных. Упрощая вопрос, можно было бы проиллюстрировать данный тезис аналогией: ключ к поиску особенностей земной атмосферы следует искать только на Земле.
В свете изложенного не вызывает сомнения, что микро- и макроэволюция представляют собой два не только гносеологически, но и феноменологически раздельных и качественно различных уровня эволюции, выделяемых из общей системно-иерархической структуры этого исторического процесса. Это означает, что к ним применима антиредукционистская концепция Ф. Энгельса о несводимости высших форм движения материи к низшим, о том, что низшие, входя в высшие, не могут определять их сущность. Запрет на поиск механизмов, лежащих вне микроэволюции, можно оценить не иначе, как неоправданную попытку ограничения свободы научного исследования, догматизации и увековечивания сложившихся теоретических стандартов. Нетрудно представить, на каком уровне остановилось бы развитие физики, а вместе с ней и производительных сил общества, если бы, скажем, Ньютон вменил в обязанность всем ученым и на все времена анализировать природные явления исключительно с позиций сформулированных им законов механики и никто из физиков последующих поколений не попытался бы нарушить этой «установки». Тогда, очевидно, у нас не было бы ни волновой теории, ни термодинамики, ни теории относительности, ни квантовой и ядерной физики.
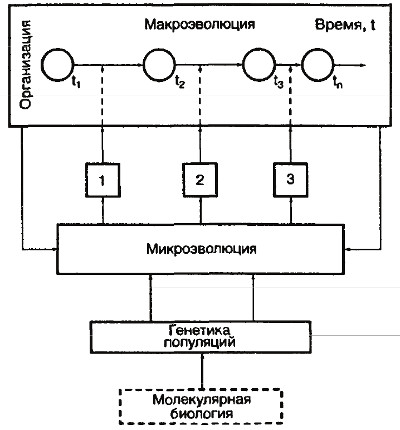
Рис. 2. Структура эволюции теории (из: Борзенков, 1987)
В заключение этого раздела целесообразно привести схему, отображающую структуру эволюционной теории по представлениям СТЭ, заимствованную из уже упоминавшейся работы В.Г. Борзенкова (1987). На схеме (рис. 2) фундамент всей теории составляет генетика популяций как единый причинный механизм эволюции. Трансформация генетической структуры популяции завершается видообразованием – центральным событием микроэволюции, очерчивающим ее верхнюю границу. Для объяснения макроэволюции в терминах микроэволюции требуется определенное число дополнительных теоретических гипотез и моделей, изображенных прямоугольниками 1–3. На схему попала и молекулярная биология, данные которой по необходимости все шире привлекаются СТЭ к трактовке микро- и макроэволюции.
Эта схема нам еще пригодится в дальнейшем, когда, опираясь на нее, мы попытаемся отобразить множественность путей макроэволюции. А пока обратимся к тому, как генетики начала XX в. мыслили себе конкретно эту надвидовую эволюцию.
