2.2. СТРУКТУРА ВИДА
Ареал. Ареал — это часть земной поверхности (территории или акватории), в пределах которой распространен данный вид. Первичный ареал соответствует области становления вида. В дальнейшем он может расширяться в результате расселения или, наоборот, сужаться вследствие вымирания. Ареал представляет неотъемлемую характеристику вида.
Ареалы вида условно подразделяются на сплошные и прерывистые (дизъюнктивные). Ареалов, сплошь заселенных особями того или иного вида, в действительности не существует. На местах, отвечающих биологическим потребностям вида, могут формироваться поселения — местные популяции. Критерию сплошного распространения соответствует регулярное нахождение таких популяций в местах, пригодных для обитания.
Примером сплошного ареала может служить ареал сибирской ели (рис. 2.1). Сибирская ель — главный образователь темнохвойного леса среднетаежной подзоны европейского северо-востока России. Она отсутствует здесь только на бесплодных песчаных почвах и на болотах. Сходную картину представляет ареал сибирской пихты, но господствует она лишь на богатых почвах, тогда как чаще встречается в виде примеси в еловых лесах. На заболоченных торфяниках повсеместно произрастают в северной лесной зоне клюква, подбел, в умеренном поясе — голубика, багульник обыкновенный.
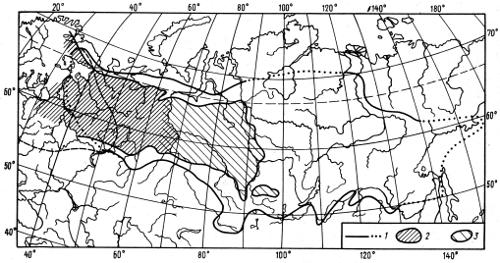
Рис. 2.1. Ареал сибирской ели Picea obovata (по А. И. Толмачеву, 1974): 1 — граница ареала, 2 — пространство, в пределах которого сибирская ель является основным лесообразователем, 3 — область согосподства ели, пихты и кедра, на севере — ели и лиственницы
Существуют виды-космополиты или полукосмополиты, ареал которых охватывает огромные пространства в биосфере Земли. К ним относятся многие морские беспозвоночные, некоторые виды плодовых мушек дрозофил, представители отряда воробьинообразных, дельфин косатка. Повсеместно распространены папоротник-орляк, обыкновенный тростник, некоторые виды рдестов.
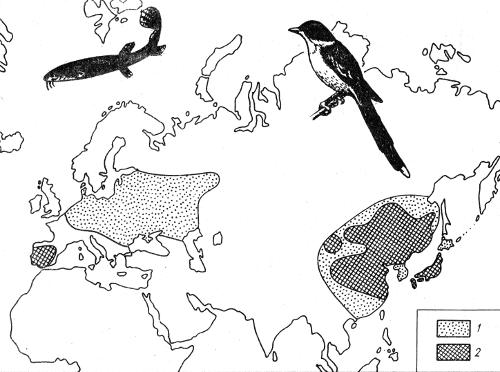
Рис. 2.2. Примеры разорванных ареалов вьюна Misgurnus fossilis (1) и голубой сороки Cyanopicia суета (2) (по А.В. Яблокову, А.Г. Юсуфову, 1989)
Для прерывистых ареалов характерны пространственные разрывы между различными частями — группами популяций, отражающие историю вида. Изменения климата в плейстоцене, сопровождавшиеся общим похолоданием, усилением суровости зим, уменьшением влажности, наиболее резко повлияли на глубинные районы Евразии, особенно на Восточную Сибирь. Здесь происходило вымирание популяций многих видов растений и животных, которое распространялось затем на всю Сибирь, а иногда и на Русскую равнину. Наступление ледников явилось основной причиной разрыва ареалов голубой сороки и вьюна (рис. 2.2), растения кислички (рис. 2.3). Теплолюбивые формы, вымершие на больших пространствах Евразии, сохранились в Средиземноморье, на Кавказе, на территории Дальнего Востока.
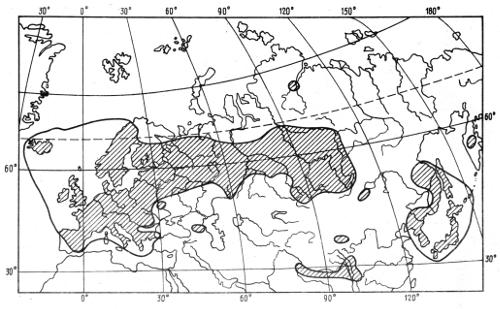
Рис. 2.3. Распространение кислички Oxalis acetosella в пределах Евразии (заштриховано) (по А.И. Толмачеву, 1974)
Узкие ареалы часто имеют островные виды. Своеобразие фауны и флоры островов выражено тем сильнее, чем дольше они оставались обособленными от основной суши. Так, на о. Мадагаскар, фауну которого выделяют даже в особую зоогеографическую подобласть, из 36 родов млекопитающих эндемичны 32 рода; из 127 родов птиц эндемична почти половина. На о. Св. Елены 85% высших растений — эндемики, на Гавайях — 82%. Понятию «островных форм» соответствуют и обитатели отдельных водоемов, горных областей, глубоких пещер. Несколько долин в горах Тянь-Шаня составляют ареал кок-сагыза, небольшой район на Балканах — ареал конского каштана. Уникальные виды рододендронов представлены в виде отдельных популяций на горе Кинабалу в Северном Борнео. Их расселение, очевидно, ограничено физико-географическими преградами. В озере Байкал обитают 250 эндемичных видов рачков-бокоплавов, 80 видов планарий, 40 видов олигохет, большое число эндемичных видов рыб. Большим своеобразием отличается фауна оз. Танганьика (в Африке), ряда других великих озер.
Крайней ограниченностью характеризуются ареалы реликтовых форм-видов с комплексом признаков давно вымерших групп. К ним относятся ящерообразная гаттерия, обитающая в глубоких норах на островах залива Пленти (Новая Зеландия); «девонская» кистеперая рыба латимерия, сохранившаяся в глубоководных участках прибрежных вод Восточной Африки; голосемянное дерево гинко, которое разводят в Китае и Японии как декоративное растение.
Многомерность (политипичность) вида. Еще Ч. Дарвин связывал процветание вида с широтой его расселения и подразделенностью на группы. Он писал: «Виды, наиболее процветающие, или, как их можно назвать, господствующие, — те, которые широко расселены, наиболее широко рассеяны по своей области и наиболее богаты особями, — чаще всего дают начало хорошо выраженным разновидностям, или, с моей точки зрения, зарождающимся видам».
Идея о дифференциации на группы внутри вида получила в дальнейшем развитие в рамках синтетической теории эволюции в виде концепции политипического вида. По Э. Майру, политипический вид можно определить как вид, состоящий из двух подвидов или более; монотипический вид на подвиды не разделяется. Политипические виды широко представлены в живой природе. На их долю, по некоторым оценкам, приходится не менее 75% от общего числа видов высших растений, птиц, млекопитающих.
Подвид — это совокупность локальных популяций, обитающих в одном из подразделений видового ареала и таксономически отличающихся от других таких популяций. Таксономические отличия признаются достаточными для выделения подвида в том случае, если не менее 75% особей, составляющих данную популяцию или группу популяций, отличаются от особей всех прочих популяций. В этом случае подвид получает наименование в соответствии с триномиальной номенклатурой (например, среднерусская лиса — Vulpes vulpes vulpes, степная лиса — V.v. stepensis). В зоологической систематике подвид — наименьшая таксономическая единица. В ботанической систематике выделяют еще экотип в качестве локальной экологической расы. Так, например, у можжевельника туркестанского описана высокогорная низкорослая разновидность Juniperus turkestanica var. fruticosa.
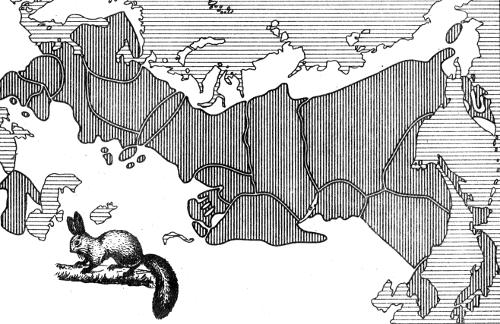
Рис. 2.4. Распределение подвидов белки Sciurus vulgaris по ареалу в Евразии (по А.В. Яблокову, Б.М. Медникову, 1986)
На рис. 2.4 показаны хорошо различимые границы подвидов белки на территории Евразии. Подвид, населяющий Кольский полуостров и Карелию, отличается частой встречаемостью краснохвостых и красноухих животных. Для белок на Украине характерно интенсивное развитие зимней коричневой окраски. Карпатские белки почти все чернохвостые и черноспинные. У якутских белок зимний мех серый, без рыжих тонов. Вообще в направлении с запада на восток увеличиваются размеры зверьков; окраска зимнего меха переходит от коричневатой к пепельно-серой и даже темной. Границы между подвидами выражены, как правило, вполне отчетливо, и их ареалы не перекрываются. Следовательно, данные подвиды являются аллопатрическими (от греч. allos — другой, patris — родина).
Примеры симпатрического (от греч. syn — вместе) существования подвидов приводит видный американский ботаник В. Грант. Однолетнее травянистое растение Gilia achilleifolia произрастает в Калифорнии, в саванне с участками виргинского дуба. Его популяции могут быть надежно отнесены к одному из двух подвидов: G. a. achilleifolia и G. a. multicaulis. Растения первого подвида широко распространены и встречаются на открытых солнечных полянах. Их крупные сине-фиолетовые цветки собраны в головки и опыляются пчелами. Растения второго подвида растут на полузакрытых участках редколесья. Они самоопылители с мелкими бледно окрашенными цветками. По всему ареалу лесные и открытые участки чередуются друг с другом и определяют тем самым характер распространения местных популяций двух подвидов (рис. 2.5).
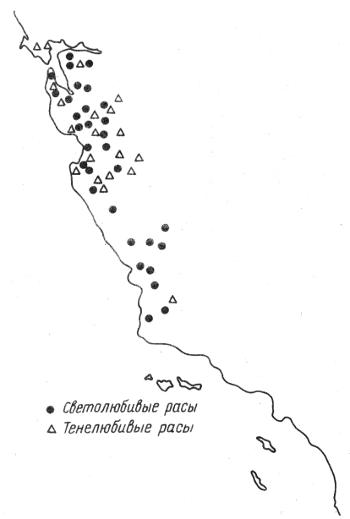
Рис. 2.5. Распространение двух экологических рас Илии Gilia achilleifolia на западе США (по В. Гранту, 1984)
Термин «экотип» введен шведским ботаником Г. Турессоном в 1922 г. Экотип представляет собой группу растений одного вида, обладающих комплексом наследственно обусловленных признаков и приспособленных к определенным условиям обитания (к определенным биотопам).
Исследования экологической дифференциации видов растений были начаты еще в начале века. Русский ученый Н.В. Цингер выявил сезонные расы, раннецветущие и позднецветущие, у большого погремка, растущего на лугах и засоряющего посевы ржи, и у рыжика, специализированного сорняка льна. Он объяснил формирование сезонных рас как результат действия отбора, направленного на выживание сорных растений в условиях покосов в летние месяцы.
Г. Турессон, Й. Клаузен, а в нашей стране Е.Н. Синская и М.А. Розанова разработали в 20—30-х годах экспериментально-экологическое направление в микросистематике, которое включает также элементы генетического анализа. Основной их метод состоял в перенесении отдельных растений или групп растений из различных частей видового ареала, контрастных по экологическим условиям, в однородную среду экспериментального поля, сада или теплицы. Если пересаженные образцы и их потомство сохраняли исходные различия в идентичных условиях разведения, их относили к разным экотипам. Синская предложила подразделять экотипы на климатические, эдафические (приуроченные к специализированным почвенным условиям) и ценотические (определяемые характером растительных сообществ). В обширных экспериментах с видами родов Atriplex, Geum, Campanula, Achillea и многих других было установлено преобладание климатических экотипов, хотя другие факторы также могли участвовать в их формировании. В большинстве случаев удалось раскрыть приспособительный смысл различий между экотипами (рис. 2.6).
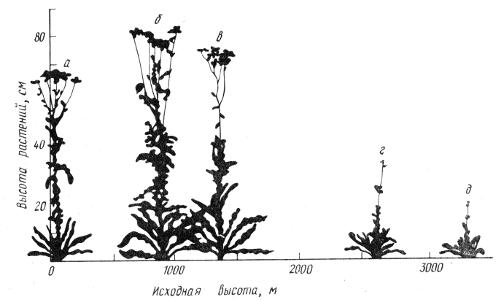
Рис. 2.6. Экотипическая дифференцировка в популяциях тысячелистника Achillea millifolium (по J. Clausen et al., 1948). Растения из мест, расположенных на разной высоте (а—д), выращивали на одном участке (высота 90 м)
Экотип Atriplex litorale, обитающий на защищенном от ветра южном побережье Швеции, представлен высокими прямостоячими растениями, тогда как экотип открытого ветрам западного побережья включает в основном низкорослые, разрастающиеся вширь растения. Для экотипов многих видов северных районов Швеции характерны более короткий вегетационный период и более ранние сроки зацветания по сравнению с экотипами южных районов.
Существование экотипов доказано и для многих видов животных, однако зоологи предпочитают применять к ним термин «экологическая раса».
«Пищевые» расы обнаружены у насекомых-фитофагов, приспособленных к разным видам растений-хозяев. Документально зафиксирован переход насекомых-монофагов к новым источникам питания. Так, яблоневая плодожорка, завезенная в Северную Америку, спустя несколько десятилетий образовала обособленные расы вредителей грецкого ореха и груши. Ивовый листоед под Санкт-Петербургом существует, как установлено, в виде ивовой и березовой рас, а в Подмосковье еще и осиновой. У хмелевого пилильщика найдены формы, связанные с хмелем и липой. Хорошо известны паразитические расы животных, растений, микроорганизмов, подразделенные по хозяину.
У видов пресноводных рыб из семейств лососевых, осетровых карповых, а также у окуней, судака имеются сезонные расы, различающиеся по срокам нереста, — так называемые «яровые» и «озимые» (Л. С. Берг). Форель, обитающая в оз. Охридском (Македония) и в оз. Севан (Армения), подразделена на экологические изоляты с характерными местами и сроками икрометания.
Биотопические предпочтения обнаруживают расы певчего воробья, серебристой чайки, дрозда-дерябы. Две формы хомячка Peromyscus polinotus, имеющие одна светлую, а другая темную окраску, приурочены соответственно к светлой песчаной почве и к темному субстрату (выходы лавы).
Вместе с тем четкая внутривидовая дифференциация не обязательно присуща даже широко расселенным видам. Такая ситуация характерна для дисперсных видов птиц, у которых молодняк мигрирует на далекие расстояния от мест выплода. Обыкновенная кряква, например, не имеет подвидов на огромном ареале Евразии и Северной Америки. Лишь на юго-востоке Гренландии обитает оседлая форма, отличающаяся сильным развитием орбито-носовых желез вследствие регулярного потребления соленой морской воды. Многие виды воробьиных птиц относятся, как показал А.С. Мальчевский, к дисперсному типу, в том числе большая синица, пеночка-весничка, дрозд-белобровик. Всех их характеризует высокая экологическая пластичность: потребление разнообразной пищи, использование различных мест для гнездования, растянутые сроки размножения.
Популяционная структура видов. Подвиды, экологические расы, экотипы складываются из местных популяций. Их число зависит от широты расселения вида, степени внутривидовой дифференциации. При изучении популяционной структуры вида прежде всего устанавливают границы между популяциями. Затем определяют степень различий между ними, характер изменчивости в последовательном их ряду. Особая задача состоит в выделении географических изолятов — популяций, обособленных от основной части видового ареала. Важное значение имеет также нахождение гибридных зон с резко повышенной изменчивостью на границах обособившихся популяций или подвидов.
Границы между популяциями выступают достаточно отчетливо, если они имеют островной (в широком смысле) характер или отделены друг от друга трудно преодолимыми естественными преградами. При нахождении границ учитывают степень подвижности животных и особенности их размножения. Некоторые летучие мыши мигрируют на зимовку за сотни и тысячи километров, но размножаются из года в год в радиусе нескольких километров. Иная картина наблюдается, как мы видели, у кряквы обыкновенной. Поэтому у таких видов выделяют репродукционный ареал — пространство, на котором происходит размножение особей; трофический ареал — площадь питания; пути миграции.
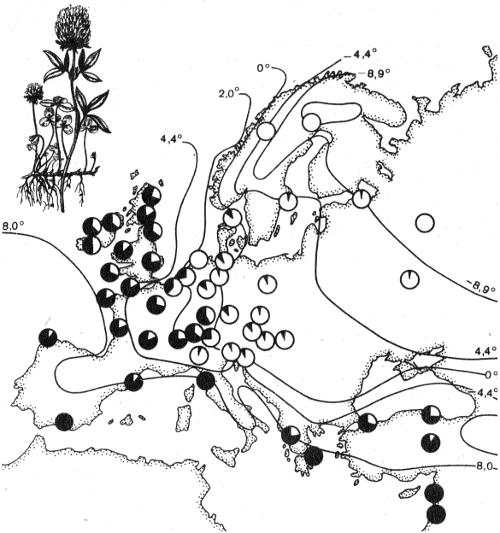
Рис. 2.7. Клинальная изменчивость содержания цианидов в белом клевере Trifolium repens, сопряженная со средней температурой января (по А.В. Яблокову, 1987): цифрами обозначены изотермы, черным сектором в кружках — доля цианидов
При наличии у вида сплошного ареала выделение популяций оказывается нередко затруднительным. Границы между ними часто бывают стертыми. Но в ряду последовательных популяций удается наблюдать постепенное изменение признаков в том или ином географическом направлении. Такой характер географической изменчивости получил название клинальной изменчивости (J. Huxley, 1939; 1942). В качестве примера рассмотрим содержание цианидов у белого клевера в ряду европейских популяций (рис. 2.7). Четко прослеживается клинальная зависимость данного признака от среднеянварских температур. Шведский энтомолог Б. Петерсен показал, что у скандинавских бабочек-капустниц из 59 изученных признаков 29 изменяются клинально. П.В. Терентьевым были выделены два подвида озерной лягушки: северная (длинноногая) и южная (коротконогая). Однако при последующем детальном анализе выборок из многих пунктов пришлось констатировать непрерывное изменение «индекса длинноногости» с севера на юг. На этом основании Терентьев отказался от первоначальной разбивки данного вида на подвиды, что вызвало критику. Потребовалось дать ответы на вопросы, означает ли стертость границ между подвидами невозможность признания их реального существования; достаточно ли градуального изменения одного признака для суждения о характере изменчивости других признаков. Необходимость решения подобных вопросов возникает каждый раз при исследовании клинальной изменчивости.
Климы (градиенты признаков) встречаются у большинства, если не у всех, континентальных видов. Их появление обусловлено постепенным изменением в пространстве климатических и прочих факторов среды, а также «потоком генов» в результате миграционных процессов. Чем ближе популяция подходит к границе видового ареала, тем сильнее сказывается необычность условий существования, пока не наступает предел дальнейшему распространению. Именно на границе ареала вида часто формируются географические изоляты. Пограничные популяции отличаются небольшими размерами, неустойчивостью, обедненностью состава.
Географический изолят определяют как популяцию или группу популяций, в которой внешние преграды препятствуют свободному обмену генами с другими популяциями данного вида. Представление о географических изолятах уже дано нами выше на примере островных форм. Можно добавить, что такие изоляты выступают как «разведчики» эволюции, поскольку они оказываются в необычных, часто экстремальных условиях. Вид ящериц Lacerta muralis на обширном пространстве Пиренейского полуострова образует три расы. В то же время на островах Балеарского и Питиузского архипелагов Средиземного моря обнаружено свыше 50 отличных друг от друга рас этого вида. Эндемичные подвиды птиц и бабочек описаны для островов оз. Виктория в Восточной Африке. На Соломоновых островах 52% видов птиц представлены изолятами. Полевка Clethrionomys glareolus у берегов Великобритании и Франции образует группу островных изолятов, которым некоторые систематики придают ранг даже самостоятельных видов. Число таких примеров можно было бы значительно умножить. К вопросу о роли изолятов в процессе видообразования и о значении гибридных зон, возникающих при вторичных контактах изолятов с родительским видом, мы еще вернемся в гл. 9.
