Механорецепция и восприятие звука
Все клетки, независимо от специализации, воспринимают механическое воздействие, в том числе и звук, но наиболее чувствительными по отношению к ним являются специализированные механорецепторные клетки. Их деятельность обеспечивает поддержание позы, координированные движения, ориентацию в пространстве, опознавание субстрата, восприятие звука, оценку давления, скоростей движения воды, ветра и т.д.
Реакция на механическое воздействие сводится в сущности к реакции на прикосновение и давление. Механические воздействия по характеру могут быть разными — постоянное давление, кратковременные деформации типа низкочастотных вибраций, звуковые и сверхзвуковые частоты. Соответственно этому существуют также различные механорецепторы. Длительное постоянное давление воспринимается рецепторами растяжения мышц, барорецепторами кровеносных сосудов, некоторыми рецепторами органов равновесия, так называемым отолитовым аппаратом. Низкие частоты улавливаются тактильными рецепторами, проприорецепторами (мышечно-суставные рецепторы); более высокие частоты (около 100 Гц) — рецепторами вибрации и органами боковой линии рыб и амфибий, и, наконец, звуковые частоты — органами слуха.
Ухо человека в норме воспринимает частоты от 15—20 Гц до 16—20 кГц. С возрастом понижается чувствительность к высоким частотам. Минимальный порог слышимости лежит при 1000—2000 Гц и составляет примерно 0,0002 дин/см2 (обычный разговор ~1 дин/см2, а звук, вызывающий болевое ощущение, например авиационные двигатели, ~300 дин/см2). Многие животные слышат звуки в более широком диапазоне частот, чем человек. Так, собаки воспринимают звуки частотой до 35 кГц, крысы и морские свинки — до 40 кГц, летучие мыши — от 30 Гц до 100 кГц, т. е. они способны слышать ультразвук (свыше 20 кГц). Это связано с тем, что летучие мыши имеют систему эхолокации. Они с помощью рта или ноздрей издают высокочастотные ультразвуковые сигналы-писки и улавливают слабое эхо, возникающее при отражении звуковых волн от удаленных объектов, и в полной темноте управляют своим полетом, избегают столкновения с предметами и ловят насекомых. Фонорецептор летучих мышей обладает очень высокой чувствительностью: он улавливает эхо, которое в 2000 раз слабее посылаемых ультразвуковых импульсов. Помимо летучих мышей, эхолокацию применяют дельфины, некоторые пещерные птицы, землеройки, водные насекомые и рыбы. Некоторые насекомые также способны воспринимать ультразвук. Так, диапазоны частот, улавливаемых кузнечиками, составляют 800—45 000 Гц, сверчками — 250—10 000 Гц, верхняя граница слышимости ночных бабочек достигает 150 кГц.
Органами слуха у некоторых насекомых являются либо простого строения хордотональные сенсиллы, выступающие из твердого покрова, либо сложного — тимпанальные органы, в состав которых может входить множество хордотональных сенсилл, укрепленных на тимпанальной мембране. Органы слуха располагаются в различных отделах тела — на лапках, на брюшке. Механорецепторные клетки, чувствительные к вибрации субстрата и воздуха, могут быть разбросаны по всему телу насекомого, но главным образом они сгруппированы в области суставов. Так, у пауков они расположены у основания каждой ноги и тонко воспринимают колебания паутины. Частота вибрации, которую воспринимают эти органы, — от 20 до 700 Гц, так что по сути дела они представляют собой низкочастотные фонорецепторы.
Все разнообразные механорецепторные клетки делятся в основном на два типа. Один из них представляет собой первично-чувствующие сенсорные нейроны с двумя отростками — центральным (аксоном) и периферическим, который оканчивается в коже или в мышечных волокнах и является чувствительным к механическим воздействиям. Иногда чувствующие нервные окончания закключены в капсулы, например в тельцах Пачини. Другой тип механорецепторных клеток представляет собой специализированные эпителиальные вторичночувствующие клетки с волосками, так называемые волосковые клетки; они имеются в боковой линии рыб, в вестибулярном и слуховом аппаратах позвоночных.
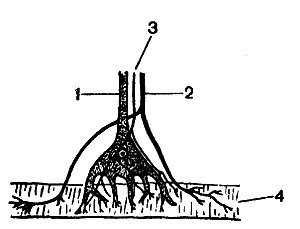
Рис. 58. Схема строения рецептора растяжения мышечных волокон рака: 1 — сенсорное волокно рецептора; 2 — моторное волокно; 3 — тормозное волокно; 4 — мышечное волокно
В качестве примера функционирования клеток первого типа рассмотрим сначала хорошо изученный рецептор растяжения ракообразных. Строение его представлено на рисунке 58. Это крупный сенсорный нейрон, многочисленные дендриты которого оканчиваются на мышечном волокне. Пассивное растяжение или активное сокращение мышцы вызывает деформацию дендритов и этим раздражает нейрон, что влечет за собой частичную деполяризацию клетки (генераторный потенциал). При достижении критического уровня деполяризации возникает импульс, бегущий но аксону. Частота сенсорных импульсов определяется главным образом величиной раздражения и скоростью его нарастания. На дендритах рецепторной клетки оканчиваются веточки тормозящего нерва; поступающие по ним импульсы из центральной нервной системы вызывают торможение, т. е. не дают деполяризации достигнуть критического уровня, при котором возникает импульс.
Подобный ход событий имеет место и при раздражении сенсорных окончаний мышечных веретен и телец Пачини. Последние обнаружены в сухожилиях, брыжейке, в коже и поджелудочной железе млекопитающих. Они достигают длины 1,0—1,5 мм и толщины 0,5 мм. Тельце Пачини состоит из многослойной наружной капсулы, внутренней колбы и нервного волокна; в пределах внутренней колбы оно не имеет миелиновой оболочки, в области наружной капсулы она есть (рис. 59). В немиелинизированном нервном окончании содержится большое количество митохондрий, главным образом поблизости от плазматической мембраны волокна в цитоплазме имеются нейропротофибриллы, ориентированные параллельно продольной оси окончания. Первый перехват Ранвье лежит внутри капсулы, а второй — вне ее. При действии давления тельце слегка изгибается, в результате чего возникает генераторный потенциал в свободном нервном окончании внутри капсулы, а затем и импульсы в перехвате Ранвье, расположенном внутри капсулы. Эффективным раздражителем для телец Пачини служит не само давление или длительность его действия, а скорость изменения механической деформации. Крупные тельца Пачини из брыжейки млекопитающих возбуждаются при смещении всего лишь на 0,5 мкм, длящемся 0,1 мс.
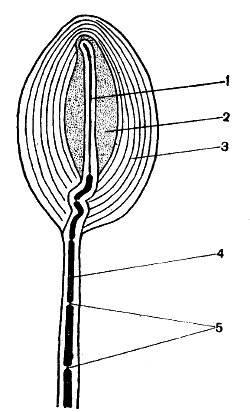
Рис. 59. Схема строения тельца Пачини: 1 — немиелинизированное нервное окончание; 2 — внутренняя колба; 3 — многослойная наружная капсула; 4 — миелинизированное нервное волокно; 5 — перехваты Ранвье
Существуют две точки зрения относительно механизма функционирования телец Пачини. Согласно одной из них (медиатор- ной гипотезе), вследствие механического воздействия структуры внутренней колбы выделяют медиатор ацетилхолин, который, в свою очередь, действует на мембрану нервного окончания и вызывает изменение ее поляризации. Согласно другой гипотезе, механическое раздражение непосредственно действует на механочувствительную плазматическую мембрану нервного окончания. Предполагается, что она обладает механочувствительными белками, изменяющими свою конформацию при механических воздействиях. В пользу этой гипотезы говорит тот факт, что поперечное сечение чувствительного нервного окончания имеет форму эллипса (отношение осей — 2,26). И если деформирующая сила действует вдоль большой продольной оси, рецепторная мембрана сжимается, так как поверхность волокна уменьшается и возникает гиперполяризация (или величина мембранного потенциала не меняется). Если сила действует вдоль малой поперечной оси, происходит растяжение мембраны, так как площадь поверхности волокна увеличивается, при этом наблюдаются деполяризация и возникновение импульсов. Импульсы возникают на каждый генераторный потенциал, вплоть до частоты раздражения 150 имп/с при слабых ударах и до 280 имп/с при сильных. Генераторный потенциал (но не нервный импульс) вызывается раздражителем вплоть до частоты 650 имп/с.
Уменьшение частоты сенсорных импульсов при продолжающемся раздражении свидетельствует об ослаблении чувствительности рецепторной клетки и представляет собой адаптацию. Некоторые механорецепторы адаптируются быстро и дают всего только один импульс на раздражение; таковы тактильные волоски и рецепторы осязания. Короткая вспышка импульсов характерна для тактильных окончаний в коже лягушки и для телец Пачини. Рецепторы растяжения в мышцах адаптируются медленнее, чем тактильные рецепторы, и могут давать разряды в течение 1 ч и более. В барорецепторах каротидного синуса млекопитающих адаптация фактически отсутствует.
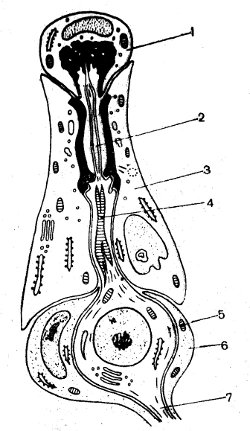
Рис. 60. Схема строения хордотональной сенсиллы тимпанального органа кузнечика (по Иванову, 1968): 1 — шапочковая клетка; 2 — видоизмененный жгутик; 3 — оберточная клетка; 4 — периферический отросток; 5 — рецепторная клетка; 6 — шванновская клетка; 7 — центральный отросток
Все механорецепторные клетки насекомых являются первично-чувствующими и имеют два отростка — центральный и периферический (видоизмененный жгутик), который и воспринимает механический стимул. На рисунке 60 представлена схема строения хордотональной сенсиллы тимпанального (слухового) органа кузнечика. Периферический, чувствующий отросток покрыт сверху шапочковой клеткой, прикрепленной к тимпанальной мембране, колебания которой являются механическим стимулом. Предполагают, что в разных участках тимпанальная мембрана обладает разной структурой, а следовательно, и разной способностью смещаться на действие звука различной частоты — тонкие и жесткие части мембраны смещаются главным образом звуками высокой частоты, а толстые и мягкие — низкими звуками. Это обусловливает локализацию восприятия разных звуковых частот. Таким образом, в тимпанальном органе насекомых, подобно кортиеву органу позвоночных, имеются низкочастотные и высокочастотные слуховые клетки.
Механорецепторные клетки органов равновесия у беспозвоночных также являются первичночувствующими клетками со жгутиками, чаще это двигающиеся киноцилии, реже Неподвижные стереоцилии. Они выстилают внутреннюю полость статоциста (орган равновесия), которая заполнена жидкостью и содержит подвижное тельце — отолит (или секретированные известковые образования или песчинки). При изменении положения тела отолит внутри пузырька перемещается и раздражает соответствующие чувствительные клетки.
Механорецепторные клетки второго типа, так называемые вторичночувствующие волосковые клетки, имеются в стенках сложной системы каналов боковой линии рыб и водных амфибий. Волосковые клетки (нейромасты) расположены в каналах группами, и каждая группа покрыта желеобразным колпачком — купулой, в канальцы которой проникают чувствительные волоски (рис. 61). Вода, свободно протекающая через поры, перемещается вдоль каналов, деформирует купулы и изгибает волоски рецепторных клеток. Нейромасты могут улавливать колебания воды с частотой не более 100 Гц. В состоянии покоя, когда давление воды со всех сторон купулы одинаково, рецепторные клетки генерируют нервные импульсы с постоянной частотой. Если возникает ток воды по каналу, частота импульсов изменяется, либо увеличивается, либо уменьшается в зависимости от направления тока воды.
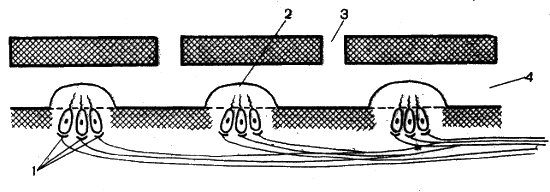
Рис. 61. Схема строения боковой линии: 1 — волосковые клетки (нейромасты); 2 — купула; 3 — пора; 4 — канал
У позвоночных волосковые клетки имеются в вестибулярном аппарате, который состоит из преддверия (отолитового органа) и системы полукружных каналов. Первый из них реагирует на линейные ускорения и отклонения от вертикали, второй — на угловые ускорения. Сенсорный эпителий с волосковыми клетками расположен на слуховых гребешках (кристах) полукружных каналов и на слуховых пятнах (макулах) отолитового органа. В кристе горизонтального канала морской свинки эпителий занимает площадь, равную 0,4 мм2, количество сенсорных клеток в нем составляет около 6000; площадь сенсорного эпителия в макуле отолитового органа равна 0,55 мм2 с 8400 сенсорными клетками. Средняя величина сенсорной клетки тритона составляет 0,024—0,030 мм.
Между волосковыми клетками расположены опорные эпителиальные клетки, имеющие в цитоплазме опорные фибриллы.
У высших позвоночных выявлены два типа сенсорных волосковых клеток. Клетки первого типа имеют форму кувшина с коротким тонким горлышком и расширенным основанием, где расположено крупное ядро. Клетки второго типа цилиндрические и более вытянутые. Различаются эти клетки не только по форме, но и по субмикроскопической организации — разный состав, расположение клеточных органелл и форма окончаний нервных волокон, вступающих в контакт с этими клетками. От клеток первого типа отходят более толстые нервные волокна, чем от клеток второго типа. Предполагают, что клетки второго типа реагируют на более сильные стимулы, а клетки первого типа — на более слабое воздействие. Оба типа сенсорных клеток несут на апикальном конце пучок чувствительных волосков (рис. 62). Он состоит из 40—70 тонких и неподвижных стереоцилий и одной длинной и подвижной киноцилии, которая всегда расположена полярно на периферии пучка. Подобно жгутику она содержит 9×2+2 фибриллы, а стереоцили — 70—80 тонких плотно упакованных фибрилл.
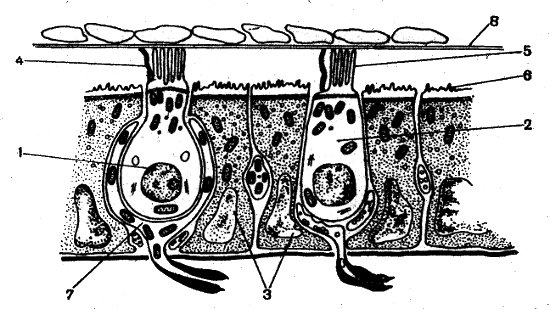
Рис. 62. Схема строения утрикулюса млекопитающих (Винников, 1966): 1 — рецепторная клетка первого типа; 2 — рецепторная клетка второго типа; 3 — опорные клетки; 4 — киноцилии; 5 — стереоцилий; 6 — микровилла; 7 — чашевидное нервное окончание; 8 — мембрана с известковыми телами (отолиты)
Сверху чувствительный эпителий кристы покрыт студенистой массой — купулой, имеющей канальцы диаметром 3—5 мкм, в которых находятся чувствительные волоски. Между кристой и купулой имеется узкое субкупулярное пространство, так что купула может смещаться относительно кристы, деформируя при этом волоски сенсорных клеток. Таким образом, всякое движение, которое возникает в эндолимфе, купула воспринимает и передает на волоски чувствительных клеток. Над сенсорным эпителием слуховых пятен вместо купулы лежит студенистая мембрана, содержащая многочисленные известковые тела (отолиты). Чувствительные волоски пронизывают эту мембрану и деформируются при ее движении. Максимальное смещение мембраны равняется 0,1 мм. В зависимости от направления смещения чувствительных волосков, наблюдается градуальная деполяризация волосковых клеток. Деполяризация, по-видимому, связана с усилением спонтанного разряда нервных импульсов, и возникает она в том случае, если механический стимул направлен к киноцилии, а гиперполяризация связана с ослаблением импульсации и возникает при действии силы в сторону от киноцилии (рис. 63). Таким образом, при раздражении может наблюдаться изменение уровня активности либо в сторону повышения, либо в сторону снижения. Это имеет значение для регулирования положения тела в пространстве.
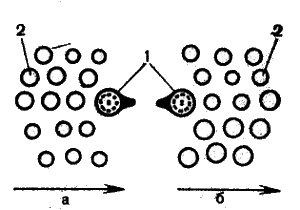
Рис. 63. Схема положения киноцилии (1) по отношению к пучку стереоцилий (2) (Lowenstein, 1964). Стрелками показано: а — возбуждающее направление наклона стереоцилий, б — тормозящее
Начальным воспринимающим звеном слухового рецептора млекопитающих являются специализированные волосковые клетки кортиева органа, расположенного в улитке внутреннего уха. Все остальные части внешнего и среднего уха служат для улавливания, концентрирования и передачи звука этим сенсорным клеткам, трансформирующим звуковые колебания в нервные импульсы. Колебания барабанной перепонки (внешнего уха) передаются посредством слуховых косточек через овальное окно эндолимфе, заполняющей внутреннее ухо, и базилярной мембране, на которой и располагаются чувствительные клетки. Их существует два вида — внутренние волосковые клетки, расположенные в один ряд (у человека их число достигает 3 500), и наружные, расположенные в три ряда (общее число 12 000—20 000). Внутренние слуховые клетки менее чувствительны.
На свободном, апикальном конце слуховых клеток имеются волоски — стереоцилий длиной 4—5 мкм и диаметром 0,35 мкм у внутренних клеток и 0,15 мкм у внешних. В плазматических мембранах стереоцилий наблюдается высокая АТФ-азная активность. Волоски контактируют с покровной (текториальной) мембраной, которая представляет собой неклеточное волокнистое образование, состоящее из гликопротеидов. При колебании базилярной мембраны волоски рецепторных клеток, касаясь покровной мембраны, деформируются, в результате чего и возбуждается слуховая клетка. У позвоночных волосковые клетки улитки возбуждаются при перемещении эндолимфы, составляющем всего лишь 10—9 см. Амплитуда и скорость колебаний различной частоты достигает максимума в различных местах базилярной мембраны. При высоких частотах возбуждается только базальный завиток улитки, а верхушка улитки наиболее чувствительна к самым низким тонам.
Соответственно этому наблюдаются и сдвиги в ультраструктурной организации волосковых клеток, особенно наружных. Происходит наклон стереоцилий, изменяются размеры ядра, наступает вакуолизация цитоплазмы, увеличивается число телец Гензена, повышается синтез белка, перераспределяются митохондрии и структуры аппарата Гольджи, в пресинаптической области наблюдается скопление синаптических пузырьков, возрастает общее количество гликогена, и он переходит из гранулированной формы в диффузную. Все это говорит о том, что реакция на звуковой стимул является тотальной, захватывающей всю клетку.
Согласно механоэлектрической теории Дэвиса, незначительная механическая деформация стереоцилий волосковых клеток, возникающая вследствие колебания основной мембраны, изменяет проницаемость и электрическое сопротивление поверхностной мембраны стереоцилий, вследствие чего возникает генераторный (микрофонный) потенциал и токи действия в нервных окончаниях, контактирующих с клеткой.
Согласно цитохимической теории Я.А. Винникова и Л.К. Титовой, движение основной и текториальной мембран приводит к проникновению ацетилхолина в область стереоцилий, где он и взаимодействует с холинрецептивным белком поверхностной мембраны стереоцилий. В результате этого взаимодействия изменяется проницаемость поверхностной мембраны стереоцилий, наступает ее деполяризация и запускается ряд ферментативных внутриклеточных реакций, приводящих к выбросу ацетилхолинового медиатора в синапсах — контактах клетки с нервными окончаниями. В последних порождаются нервные импульсы, поступающие в слуховые центры.
Несмотря на разнообразие механорецепторов, принцип действия у них один — деформация высокочувствительных к механическим воздействиям структур вызывает градуальную деполяризацию клеточной мембраны с последующим возникновением генераторного потенциала и нервных импульсов. Как совершается трансформация механической энергии в электрическую, т. е. какие события совершаются между деформацией и рождением нервных импульсов, пока еще не ясно. Несомненно лишь, что это преобразование механической энергии в электрическую сопровождается не только событиями на поверхностной мембране, но и внутри клетки и связано с процессом обмена веществ.
