Фоторецепция
Все клетки способны реагировать на свет, но те из них, которые содержат пигмент, более чувствительны к действию света, Если искусственно ввести краситель в клетку, то также наблюдается повышение ее светочувствительности (фотодинамический эффект).
С наличием желтого пигмента растений — каротиноида — связывают фототропизм растений, т. е. изгибание растения в сторону источника света. Особенно хорошо это проявляется в синем свете. Микроорганизмы — пурпурные бактерии, сине-зеленые и зеленые жгутиковые водоросли — реагируют на свет разнообразными фототаксисами; полагают, что в рецепции света принимают участие флавины и каротиноиды. Световой чувствительностью обладают клетки радужной оболочки глаза, пигментированные нейроны змей, речного рака и гигантские нервные клетки брюхоногого моллюска (Aplysia), имеющие пигменты гемопротеиды и каротинпротеиды.
Непосредственно на свет могут реагировать и хроматофоры — специализированные пигментные клетки покровных тканей, обеспечивающие маскирующую защитную окраску животного, терморегуляцию, защиту от интенсивного освещения и брачную расцветку. Хроматофоры обнаружены у рептилий, земноводных, рыб, круглоротых, ракообразных, насекомых, головоногих моллюсков, аннелид и иглокожих. Хроматофоры различаются по форме, типу пигмента и реакциям на раздражение. У ракообразных, насекомых и рыб хроматофоры содержат два или даже более пигментов, у амфибий и рептилий, как правило, один пигмент, часто меланин.
Хроматофоры способны содержащиеся в них зернышки пигмента либо концентрировать в маленький шарик в центре клетки, что приводит к посветлению окраски, либо диспергировать их по всей клетке, обычно сильно разветвленной, что вызывает потемнение окраски животного (рис. 51).
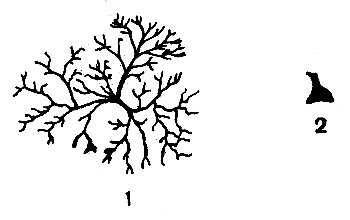
Рис. 51. Хроматофор десятиногого рака (Palaemonetes): 1 — пигмент диспергирован; 2 — пигмент сконцентрирован
При диспергировании пигмента его количество может увеличиваться за счет вновь образующегося, в концентрированном же состоянии, наоборот, количество пигмента может уменьшаться за счет его разрушения. Эти реакции хроматофоров, часто чрезвычайно сложные, в норме регулируются эндокринной и нервной системами с участием рецепторов, в первую очередь зрительных. Хроматофоры и сами по себе способны непосредственно реагировать на свет, но эта первичная реакция в условиях целого организма маскируется и не проявляется вследствие гораздо более активных регулирующих нейрогуморальных влияний.
Первичную реакцию хроматофоров холоднокровных животных в чистом виде можно наблюдать на изолированных кусочках кожи или после полной денервации пигментных клеток и удаления гипофиза (в нем вырабатываются гормоны, регулирующие деятельность хроматофоров). У молодых особей амфибий и рыб, хроматофоры которых еще не находятся под контролем регуляторных систем организма, также проявляется непосредственное действие света на пигментные клетки: животные темнеют на свету и светлеют в темноте.
Специализированные светочувствительные клетки органов зрения — фоторецепторы — содержат пигмент, называемый зрительным. По структуре и функциональным возможностям органы ярения очень разнообразны — от простых одноклеточных рецепторов кольчатых червей и глазков кишечнополостных, способных различать лишь интенсивность света, до сложных фасеточных глаз насекомых и камерных глаз позвоночных, которые уже обеспечивают образное зрение. В органах зрения наряду с собственно рецепторными клетками существуют вспомогательные структуры, регулирующие освещение чувствительных клеток.
Рассмотрим строение наиболее изученных светочувствительных клеток сетчатки глаза позвоночных. Последние обладают двумя типами фоторецепторных клеток — палочками и колбочками. Палочки гораздо чувствительнее колбочек, они воспринимают свет слабой интенсивности, имеют малую разрешающую способность и не различают цвета, т. е. являются рецепторами сумеречного зрения. Колбочки — аппарат дневного и цветового зрения, они функционируют лишь при большой интенсивности света, обладают слабой световой чувствительностью, но большой разрешающей способностью. Поэтому у ночных животных, как правило, в сетчатке преобладают палочки, а у дневных — колбочки. Колбочки быстро адаптируются в темноте, палочки — медленно.
Величина фоторецепторных клеток различна у разных животных. У лягушки, например, длина палочки составляет 55 мкм, а диаметр — 5—6 мкм; у быка длина палочки составляет лишь 10 мкм, а диаметр — 1 мкм.
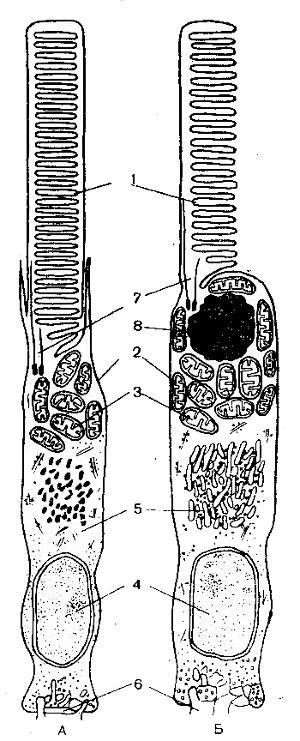
Рис. 52. Схема строения палочки (А) и колбочки (Б) (Говардский и Грибакин, 1975): 1 — наружный сегмент; 2 — внутренний сегмент; 3 — эллипсоид (скопление митохондрий); 4 — ядро; 5 — комплекс Гольджи; в — синаптическое тельце; 7 — цилия (соединительная ножка); 8 — масляная капля
По внешнему виду палочки и колбочки похожи друг на друга, только палочки уже колбочек. На рисунке 52 представлено схематическое изображение палочки и колбочки. Эти клетки состоят из наружного сегмента, содержащего зрительный пигмент, внутреннего сегмента с митохондриями, ядерно-плазматического и синаптического телец, куда подходят дендриты нейрона. Наружный сегмент представляет собой видоизмененный жгутик и содержит стопку дисков (толщиной 14 нм), состоящих из, сдвоенных мембран. Последние образуются из поверхностной плазматической мембраны жгутика; в колбочках они остаются связанными с нею, а в палочках диски отрываются от поверхностной мембраны и превращаются в замкнутые структуры. Число дисков различно у разных животных. Так, палочки у лягушки содержат 1100 дисков, а палочки быка — 180 дисков.
В этих мембранных структурах и расположен зрительный пигмент, состоящий из хромофорной группы и белка опсина. Пигмент составляет значительную часть наружного сегмента — 80—95% от всего белка фоторецепторной мембраны. Помимо белков, в состав мембраны входят липиды, главным образом фосфолипиды. Предполагают, что молекулы зрительного пигмента частично погружены в липидный слой и свободно перемещаются в плоскости диска по типу броуновского движения. Здесь, на фоторецепторной мембране, и совершается первичный акт световой рецепции — молекулярные превращения и распад зрительного пигмента под действием света.
Наружный сегмент прикреплен к внутреннему тонкой ножкой (цилия). В результате сокращения фибрилл ножки, а также наружного и внутреннего сегмента, по-видимому, и осуществляются ретиномоторные явления — сокращение и удлинение палочек и колбочек. На свету палочки вытянуты, колбочки сокращены; в темноте, наоборот, палочки сокращены, колбочки вытянуты. При ретиномоторных явлениях расстояние между мембранами наружного сегмента изменяется, что связано с их функционированием и адаптацией к свету и темноте.
В верхней части внутреннего сегмента имеется скопление крупных вытянутых и иногда ориентированных вдоль клетки митохондрий (образующих так называемый эллипсоид), что говорит об интенсивной синтетической деятельности клетки. При помощи метода авторадиографии с использованием меченых аминокислот было показано, что белки фоторецепторных клеток, в том числе и опсин, синтезируются в рибосомах внутреннего сегмента в области так называемого миоида, где расположены эндоплазматическая сеть и комплекс Гольджи. Отсюда белок расходится по всей клетке, но основная его доля мигрирует через цилию в базальную часть наружного сегмента, где и образуются мембраны дисков у палочек. В палочках лягушек в среднем в течение часа образуется около 1,5 диска. Эти мембраны, содержащие зрительный пигмент, постепенно продвигаются от базального к апикальному концу наружного сегмента и приблизительно через 8,5 недели разрушаются и фагоцитируются соседними клетками пигментного эпителия сетчатки. Освободившаяся хромофорная группа вновь возвращается в наружный сегмент и используется здесь для построения новых молекул зрительного пигмента. Благодаря наличию в клетках пигментного эпителия микроворсинок увеличивается поверхность их соприкосновения с фоторецепторными клетками. Таким образом, между фоторецепторными и пигментными клетками существует тесная функциональная связь. Помимо этого пигментные клетки имеют длинные отростки, которые располагаются между наружными члениками фоторецепторов. На свету гранулы пигмента фусцина находятся в отростках и этим экранируют фоторецепторные клетки, в темноте они перемещаются из отростков в надъядерную область.
В фоторецепторных клетках птиц, пресмыкающихся, амфибий и ганоидных рыб в апикальной части внутреннего сегмента имеется еще масляная капля красного, желтого Или желто-зеленого цвета. Видимо, она играет роль дополнительного светофильтра.
Фоторецепторная мембрана светочувствительных или ретинулярных клеток беспозвоночных (насекомые и головоногие) организована в плотно упакованные пальцеобразные выросты — микровилли, которые образуют так называемый рабдомер, являющийся аналогом внешнего сегмента фоторецепторных клеток позвоночных. Небольшое число ретинулярных клеток (обычно восемь), группируясь вместе, образуют ретинулу, а их рабдомеры объединяются в единую структуру — рабдом.
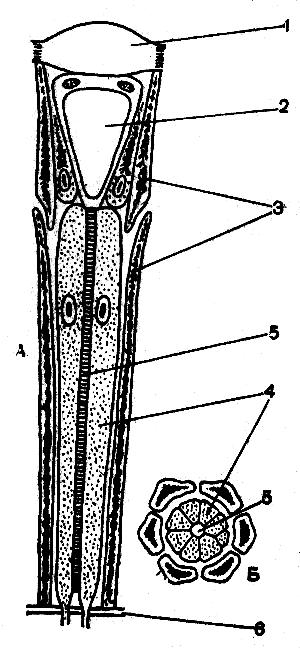
Рис. 53. Схема строения омматидия глаза пчелы (Мазохин — Поршняков, 1965). А — продольный разрез; Б —поперечный разрез; 1 — хрусталик; 2 — кристаллический конус; 3 — пигментные клетки; 4 — зрительные клетки; 5 — рабдом; 6 — базальная мембрана
На рисунке 53 изображено продольное и поперечное сечения отдельного глазка — омматидия пчелы. Пигментные клетки располагаются между светочувствительными клетками и перемещаются в соответствии с состоянием световой или темновой адаптации.
В ретинуле пчелы имеются три типа ретинулярных клеток, различающихся по размерам, глубине расположения ядра, по диаметру микровилл и восприятию разных длин излучения, что обусловливает цветовое зрение пчел. Диапазон цветов, воспринимаемых насекомыми, отличается от диапазона высших позвоночных. Насекомые видят ультрафиолетовый свет, но не воспринимают красный; они способны определять положение плоскости поляризации света, что и используется ими для ориентации в пространстве.
Помимо насекомых, цветовое зрение есть у крабов, креветок, каракатицы, у многих костистых рыб, рептилий, птиц. Среди млекопитающих цветовое зрение твердо установлено для приматов. Цветовое зрение у позвоночных обеспечивается колбочками, благодаря тому что они содержат три пигмента, поглощающих красный, зеленый или синий цвет; отсутствие одного из пигментов ведет к цветовой слепоте.
Механизм цветового зрения до сих пор остается неизвестным; ясно только, что обеспечивается оно не только периферическим зрением — зрительными пигментами, но в нем принимают участие и центральные компоненты зрительной системы.
Все известные зрительные пигменты имеют один план строения и состоят из белка опсина и хромофорной группы, которая представляет собой либо ретиналь, либо 3-дегидроретиналь (альдегиды витаминов A1 и A2). У разных видов животных зрительные пигменты варьируют (родопсин, порфиропсин, иодопсин, цианопсин и др.) за счет изменения белка опсина, а не хромофорной группы.
В темноте опсин и хромофор связаны друг с другом, при этом последний находится в определенной сложенной геометрической конфигурации (цисформа), и молекула его всегда ориентирована в плоскости, перпендикулярной направлению светового луча, что обеспечивает максимальное поглощение света. На свету же ретиналь (3-дегидроретиналь) переходит в выпрямленную (транс-форму) и разрывается связь с опсином; зрительный пигмент при этом обесцвечивается. Распад его проходит через промежуточные, неустойчивые при комнатной температуре стадии.
При освещении также меняется конформация белка опсина, молекула его становится более развернутой, уменьшается количество спирализованных участков (с 60 до 40%) с освобождением сульфгидрильных групп и изменением изоэлектрической точки. Перестройка затрагивает и липиды мембраны. Имеется представление о том, что обесцвечивание пигмента сопровождается более глубоким погружением молекулы родопсина в липидный слой. Изменяется и структура фоторецепторной мембраны, ее поверхность уменьшается, а при очень интенсивном освещении может произойти полный распад мембраны.
В темноте зрительный пигмент с помощью ферментов вновь воссоединяется, трансформа ретиналя превращается в цисформу и присоединяется к опсину. Во время световой адаптации концентрация зрительного пигмента уменьшается, а во время темновой адаптации пигмент восстанавливается и чувствительность к свету растет. При умеренном освещении устанавливается равновесие между распадом и синтезом пигмента.
Таким образом, световой сигнал трансформируется в химический благодаря молекулярным превращениям зрительного пигмента, которые являются пусковым механизмом возбуждения всей фоторецепторной клетки: происходит округление и набухание митохондрий, увеличивается количество ядерной РНК, наблюдаются цитохимические сдвиги, связанные с обменом гликогена, снижается число гранул гликогена, возникает рецепторный потенциал, который для фоторецепторов оказался гиперполяризацией. Это связано с тем, что свет уменьшает проницаемость мембраны наружного сегмента для ионов натрия и снижает вследствие этого так называемый темновой ток (который выходит из тела клетки и внутреннего сегмента и входит в наружный сегмент, замыкаясь через цитоплазму рецептора), что эквивалентно появлению фототока, выходящего из наружного сегмента и входящего во внутренний. Возникающая при освещении гиперполяризация наружного сегмента длится в течение всего периода освещения и сопровождается повышением сопротивления поверхностной мембраны на 20—50%. Таким образом, зависящая от света натриевая проводимость принадлежит лишь мембране наружного сегмента. В результате возбуждения клетки возникают нервные импульсы, поступающие в центральную нервную систему.
