Потенциал действия
В проводящих тканях (нервные, мышечные волокна, некоторые растительные клетки) помимо ПП существует так называемый потенциал действия (ПД), который сопровождает бегущую волну возбуждения и является ее рабочим механизмом, обусловливающим ее движение по волокну, иногда на большие расстояния (до 1,5 м у взрослого человека). Величина ПД при распространении его по волокну остается постоянной. Существует четко выраженный порог силы раздражения для вызова ПД, причем величина последнего не зависит от силы раздражения (сверхпороговой). Это положение известно как закон «все или ничего». В экспериментальных условиях ПД чаще всего вызывают, раздражая волокно электрическим током. ПД возникает под катодом (выходящий электрический ток), когда деполяризация волокна достигнет критического уровня. При внутриклеточном отведении ПД имеет вид, изображенный на рисунке 31. При введении микроэлектрода в волокно сразу регистрируется МП (в данном случае около 80 мВ, E1). При возникновении ПД происходит крутое падение МП до нуля, а затем он приобретает противоположный знак — плюс оказывается на внутренней стороне мембраны. Эта разность потенциалов, называемая «овершут», может достигать + 40 мВ (E2), так что величина ПД оказывается равной в приведенном на рисунке случае 120 мВ (80+40). Восходящая фаза ПД протекает очень быстро (около 0,5 мс), нисходящая фаза является гораздо более длительной и может включать в себя медленные, длящиеся десятками миллисекунд, следовые (негативные и позитивные), низковольтные потенциалы. Высоковольтную часть ПД принято называть спайком. Величину ПД можно вычислить также по формуле Гольдмана, но коэффициент проницаемости PNa+ будет равен 20 (вместо 0,04). Это связано с тем, что в момент восходящей ветви тока действия в возбужденном участке волокна проницаемость для ионов натрия резко возрастает (в 500 раз), они поступают в волокно и перезаряжают мембрану.
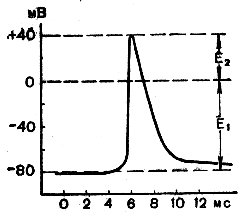
Рис. 31. Потенциал действия мышечного волокна лягушки (Nastik a Hadgkin, 1950). По оси абсцисс — время, в миллисекундах; по оси ординат — разность потенциала, в мВ
Протекание спайка сопровождается увеличением электропроводности мембраны, ее сопротивление падает с 1000 Ом/см2 в состоянии покоя до 25 Ом/см2 в активном состоянии. После кратковременной (1—2 мс) повышенной проницаемости мембраны для ионов натрия наступает повышенная проницаемость ее для ионов калия, и они по градиенту концентрации выходят из клетки наружу и создают ПД на мембране, равную равновесному калиевому потенциалу с отрицательным знаком на внутренней стороне мембраны, и тем самым снижают спайк (рис. 32). Таким образом при каждом импульсе в волокне увеличивается содержание ионов натрия и снижается количество ионов калия. Эта разница ликвидируется в последующем периоде восстановления. Изложенная натриевая теория потенциала действия была разработана английским ученым Алланом Хочкиным с сотрудниками. Потенциалы действия некоторых гладкомышечных клеток обусловлены ионами кальция, а не натрия.
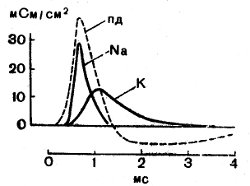
Рис. 32. Кривые проводимости для Na+ и К+и вычисленный ПД. По оси абсцисс — время, в мс; по оси ординат — проводимость, в мс (миллисименсах.) (Ходжкин, 1958)
В настоящее время накапливаются экспериментальные данные, указывающие на то, что при проведении тока действия происходят структурные изменения поверхностной мембраны. Они зависят от напряжения на мембране и сопровождаются перераспределением зарядов на мембране и появлением так называемого воротного тока или тока смещения, который и обусловливает увеличение натриевой проницаемости мембраны при ее деполяризации. Величина воротного тока гораздо меньше, чем ионные токи.
В таблице 13 представлены величины ПД для ряда объектов. Следует подчеркнуть, что они, как правило, превышают величину МП (сравните с таблицей 12).
| Таблица 13. Потенциал действия (внутриклеточные отведения) | |
| Объект | ПД, мВ |
| Аксон кальмара | 90 |
| Нерв краба | 120 |
| Мышечное скелетное волокно лягушки | 130 |
| Сердечная мышца собаки | 120 |
Длительность ПД для разных объектов варьирует от 0,6 мс до 20 с (табл. 14). Длина электрической волны довольно велика, на мышечном волокне лягушки и. нервном волокне краба она составляет 1—3 мм.
| Таблица 14. Длительность потенциала действия | |
| Объект | мс |
| Мякотное нервное волокно кошки | 0,6 |
| Безмякотное нервное волокно кошки | 10,0 |
| Нерв краба | 0,8—1,0 |
| Мышечное волокно лягушки (портняжная мышца) | 4,0 |
| Запиратель клешни краба | 150—300 |
| Мышца ноги моллюска мидии | 1000—2800 |
| Водоросль нитчатка | 1000—10 000 |
| Водоросль хара | 20 000 |
Скорость проведения ПД варьирует в зависимости от вида волокна и от его диаметра; волокна с большим диаметром проводят быстрее, чем тонкие; мякотные волокна (покрытые миелиновой оболочкой) проводят быстрее безмякотных (табл. 15).
| Таблица 15. Скорость проведения потенциалов действия | |
| Объект | мс |
| Нервные двигательные волокна | |
| человека | 60—120 |
| лягушки | 30—47 |
| рака | 6—12 |
| моллюска | 0,4—4,0 |
| Мышечные скелетные волокна | |
| кошки | 6 |
| лягушки | 2—4 |
| Растительные клетки | |
| нитчатки | 0,02 |
| мимозы | 0,05 |
В основе распространения ПД по волокну лежит местный (локальный) ток, который течет между отрицательно заряженным возбужденным участком и соседним, покоящимся (рис. 33, I). В возбужденном участке токи входят в волокно (подобно аноду) и успокаивают его (повышают МП), а в покоящемся участке токи выходят из него (подобно катоду), т. е. его деполяризуют и возбуждают. Этот местный ток является достаточным, чтобы снизить МП до критического уровня и этим вызвать ПД в соседнем участке, и так далее от точки к точке по всему нервному волокну, так что продвижение ПД происходит непрерывно.
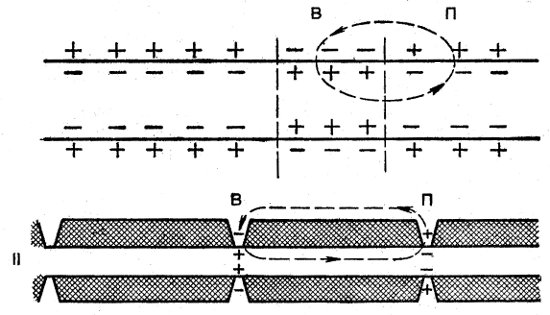
Рис. 33. Проведение потенциала действия в безмякотном (I) и мякотном (II) волокне: В — возбужденный участок; П — покоящийся участок
В миелинизированных нервных волокнах продвижение ПД по волокну происходит скачкообразно от одного перехвата Ранвье к другому (рис. 33, II). Это связано с тем, что сопротивление в перехвате Ранвье гораздо меньше, чем сопротивление миелиновой оболочки. Такой скачкообразный тип проведения импульса называется сальтаторным, он выгодно отличается от сплошного по скорости и по энергетическим затратам.
Таким образом, бегущий потенциал действия служит для быстрой передачи сигналов на относительно большие расстояния. Распространение его по волокну с постоянной величиной, без декремента, обеспечивает функцию проведения. Нервные волокна могут пропускать огромное количество импульсов (более 100 000) без видимого утомления с максимальной частотой 500—1000 имп/с. В нормальных условиях нервные импульсы передаются, как правило, с частотой 5—100 имп/с.
Важную функциональную роль играют также биопотенциалы, возникающие в специальных электрических органах, имеющихся у некоторых групп пресноводных и морских рыб; они являются органами защиты и нападения. Рыбы, живущие в мутной воде, с помощью электрических органов ориентируются в среде и избегают препятствия. Это им удается благодаря электроэхолокации. Электрические органы генерируют слабые токи; электрические поля, возникающие вокруг, искажаются любыми предметами, встречающимися на пути рыбы; последние же способны воспринимать свои слабые электрические поля и чувствовать их искажения. Некоторые морские угри через импульсную электрическую сигнализацию весьма избирательно связываются друг с другом.
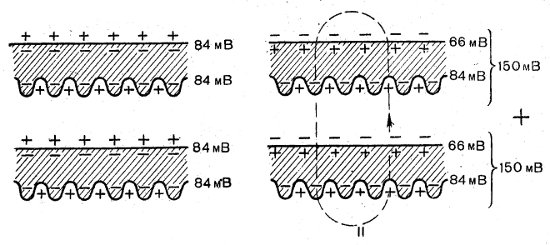
Рис. 34. Схема пары электропластинок электрического угря в покое (I) и во время спайки (II), показывающая суммирование потенциалов последовательно соединенных пластинок
Электрический орган состоит из столбиков пластинок-дисков. У ската торпедо каждый орган содержит примерно 45 столбиков по 400 пластинок; у электрического угря — около 70 столбиков по 6000 пластинок. Электропластинки соединены последовательно, а столбики между собой параллельно. Электропластинка представляет собой многоядерную распластанную клетку. Одна ее сторона гладкая, иннервированная; другая без иннервации, часто бывает складчатой. Величина МП пластинки угря равняется 90 мВ. При разрядке иннервируемая поверхность меняет полярность на обратную в 60 мВ, а потенциал складчатой поверхности не изменяется. Таким образом, две последовательно соединенные поверхности дают общий клеточный потенциал в 150 мВ (рис. 34). Напряжение, развиваемое электрическими органами, объясняется суммацией разностей потенциалов, возникающих в отдельных последовательно соединенных клетках в результате их асимметричной деполяризации. Электрические разряды отдельных пластинок строго синхронизированы, что способствует получению максимальной выходной мощности. Развиваемое электрическим органом напряжение достигает иногда 600—800 В.
