19.3. Эволюция островных экосистем. Динамика видового состава экосистем
Изменение состава экосистем в прошлом (особенно при сильных климатических возмущениях) и в настоящем связано с вымиранием одних видов и иммиграцией других. На модели экосистем островов разной величины показано (Р. МакАртур, Э. Уильсон), что вымирание видов на маленьких островах компенсируется появлением новых иммигрантов и скорость вымирания имеет тенденцию возрастания с увеличением числа видов (рис. 19.4). Число иммигрантов на островах пропорционально уменьшалось по мере увеличения общего числа видов в сообществах, как результат усиления межвидовых взаимодействий и повышения стабильности экосистемы. Равновесие сообществ было более устойчивым на островах большего размера и при наличии большого числа компонентов. Эти выводы были подтверждены на примере изучения сообществ Британских островов, на Новой Гвинее и Тринидаде, а также сообществ горных цепей. Здесь уместно вспомнить заключение Ч. Дарвина о большей интенсивности процесса «фабрикации» видов на крупных островах.
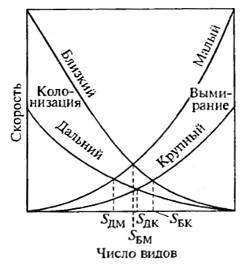
Рис. 19.4. Модель взаимодействия изоляции и площади местообитания, показывающая возможность существования нескольких равновесных состояний в процессе проникновения новых видов и темпа вымирания аборигенных форм. Равновесное число видов при прочих равных условиях будет меньше на удаленных островах, на более мелких островах и на островах с менее разнообразными биотопами, SДМ, SБМ, SДК, SБК — точки равновесия (по Р. МакАртуру и Е.О. Вильсону, 1967)
Несмотря на достигнутые успехи в изучении закономерностей динамики экосистем, все еще представляется трудной задача предсказания видового состава будущих сообществ. Это пока решается с множеством оговорок, исходя только из возможного влияния ближайших сообществ. Видовой состав будущих сообществ зависит также от эволюционной истории. Так, дятлы как насекомоядные представляют компонент многих сообществ. Однако их нет на Галапагосских островах, хотя там много насекомых; их функцию там выполняют некоторые вьюрки, клюв которых короче клюва дятла. Для добычи насекомых из трещин коры деревьев вьюрки пользуются колючками кактусов (держа их в клюве, они выковыривают насекомых из-под коры). Точно так же экологическую роль плодоядных обезьян в лесах Азии и Новой Гвинеи выполняют птицы.
Трудно предсказуемы и различия в числе видов, обитающих примерно на одинаковой площади в разных местах: есть попытки связывать их с совпадением условий для конкуренции видов, наличием в составе сообществ видов, подверженных резким перепадам численности, возрастом местообитания и климатом. По мере развития сообщества оно насыщается большим числом видов, что приводит к усложнению межвидовых отношений и увеличению числа действующих в нем биотических факторов. Увеличение числа видов в сообществе способствует более или менее равномерному распределению экологических факторов между видами, т. е. повышению стабильности сообщества. Этим отчасти и обусловлена устойчивость многокомпонентных сообществ. Разрушение таких сообществ также происходит, и процесс их развития повторяется, что показано на примере сообществ рек после выброса токсических веществ (химикатов) и полей после неумелого применения удобрений. Подобные стрессовые воздействия влекут за собой усиление действия одного из многочисленных экологических факторов и доминирование небольшого числа наиболее устойчивых видов.
Обнаружена и другая тенденция в развитии экосистем: преобладание в ее составе мелких животных над крупными. Замечено, что с увеличением размера тела особей втрое численность вида уменьшается в 10 раз. Полагают, что это связано с ослаблением ожесточенности борьбы за места обитания у мелких животных по сравнению с крупными; мелкие животные легче уживаются на ограниченных площадях.
