15.5. Темпы эволюции групп
Эволюция как грандиозный процесс изменения органического мира Земли характеризуется определенными временными параметрами. Скорость (темп) — одна из самых важных особенностей процесса эволюции в целом. Выяснение конкретной скорости эволюционного изменения групп часто важно и для решения чисто практических задач, например, связанных с возникновением и распространением новых болезнетворных микроорганизмов, появлением резистентности насекомых к инсектицидам и др.
В эволюции большее значение имеет не абсолютное, астрономическое, время, а число поколений. Причины этого будут понятны, если вспомнить, что механизм возникновения адаптаций сводится, по существу, к изменению концентрации аллелей в популяции в чреде поколений (см. гл. 11). Однако для многих ископаемых форм мы не можем с достаточной точностью оценивать скорость их эволюции числом поколений. Поэтому, несмотря на некорректность астрономического времени для характеристики темпов эволюции, приходится пользоваться и этим параметром.
Существует и другой подход для определения темпа эволюции группы — учет скорости смены видов (точнее — фратрий) в одном филогенетическом ряду (Дж. Симпсон).
Проблема выбора критериев скорости эволюции. Определить скорость эволюции не всегда просто. При рассмотрении темпов эволюции часто имеют в виду изменение не только целых групп, но и отдельных признаков. Хотя изменения групп основаны на онтогенетических дифференцировках (на появлении и развитии отдельных признаков в онтогенезе особей, см. гл. 14), все же эти проблемы различны; проблема скорости изменения отдельных признаков должна рассматриваться вместе с другими особенностями эволюции органов и функций (см. гл. 16).
Теоретически скорость эволюции вида можно определить по темпу его изменения в ходе филетической эволюции. Но как измерить этот темп? Вероятно, таким мерилом могло бы быть число элементарных эволюционных явлений (см. гл. 8) в расчете на поколение, а затем число возникших за определенное время (число поколений) адаптаций. Однако пока практически определить названные параметры вряд ли возможно для любой группы. Гораздо удобнее определить темп эволюции группы по числу возникающих видов за определенное время.
Темпы формообразования. Материал, накопленный в настоящее время в ботанике и зоологии, позволяет установить два разных по времени способа видообразования: «внезапный» и «постепенный». Рассмотрим несколько примеров.
Внезапное видообразование связано с быстрой перестройкой всего генома, например при полиплоидии. Известно, что возникновение полиплоидного организма происходит в течение считанных минут. Возникшая полиплоидная особь генетически изолирована от всех остальных особей вида. Часто полиплоидные особи оказываются более устойчивыми по отношению к действию экстремальных факторов внешней среды и получают широкое распространение в условиях, где родительская форма не выживает. Таким образом, новые особи, репродуктивно изолированные от остальных особей в популяции, могут возникнуть за одно поколение. Такой процесс мог лежать в основе возникновения полиплоидных видов в разных группах. Полиплоидные ряды видов известны для многих растений (см. рис. 6.28 и 13.5). Картофель, белый клевер, люцерна, тимофеевка и другие полиплоидные растения возникли таким путем.
Другой тип полиплоидии связан с гибридизацией (аллополиплоидия), объединением геномов сравнительно близких видов (см. гл. 6 и 13). Г.Д. Карпеченко путем гибридизации капусты и редьки была синтезирована новая форма — редько-капуста (Raphanobrassica) (рис. 15.10). Б.Л. Астаурову удалось получить аллодиплоид двух видов шелкопряда: Bombux тоri и В. mandarina.

Рис. 15.10. Схема синтеза формы видового ранга — редько-капусты; А — стручок редьки Raphanus sativus и ее хромосомный набор (18R); Б — стручок капусты Brassica oleracea и ее хромосомный набор (18B); В — первичный, полностью стерильный из-за нарушения мейоза гибрид с хромосомным набором 9R + 9B; Г— аллотетраплоид с хромосомным набором 18R + 18B вполне плодовитый, так как в мейозе 18 хромосом R конъюгируют друг с другом и образуют 9 бивалентов, как и 18 хромосом В (по Г.Д. Карпеченко, 1935)
Наконец, причиной быстрого изменения генома, определяющего генетическую изоляцию вновь возникшей особи от исходной формы, могут быть перестройки хромосом (фрагментация и слияние). У растений возможность участия межхромосомных перестроек в эволюции была экспериментально подтверждена созданием скерды (Crepis nova), отличающейся по морфологии хромосом от родительского вида. У животных роль хромосомных перестроек в видообразовании была показана в обширной серии экспериментальных и популяционно-генетических работ в группе видов дрозофилы Drosophila persimilis — virilis (см. гл. 6, рис. 6.27).
Следовательно, в природе возможно быстрое (практически внезапное) образование особи, репродуктивно изолированной от остальных особей того же поколения. В некоторых редких случаях такие особи оказываются способными к размножению и на протяжении последующих поколений могут дать начало целой группе подобных особей. Естественно, что одна-единственная особь (или даже несколько особей) видом названа быть не может уже хотя бы потому, что всякий вид — это всегда генетическая система с множеством свойств, присущих именно системе особей, в том числе с собственным ареалом, многомерной экологической нишей и т. д. Получается, что в природе могут существовать особи, уже не относящиеся к старому виду и еще не образовавшие нового вида.
Решение этой проблемы нашел К.М. Завадский, предложивший называть такие новые формы не видами, а формами видового ранга. Ясно, что из многих таких «кандидатов» в новые виды лишь небольшое число действительно превратится в самостоятельный вид. Для этого они должны оказаться достаточно жизнеспособными и иметь преимущества перед родительскими формами, позволяющими им распространиться и занять определенный ареал. Такое превращение формы видового ранга (представленной сначала немногими особями, а потом отдельными мелкими группами) в реальный вид должно продолжаться значительное число поколений и осуществляется всегда под контролем естественного отбора.
Более обычно в эволюции постепенное формообразование. Сейчас накопилось много данных, позволяющих сравнительно точно определять скорость образования подвидов у самых разных групп. Многие из них касаются послеледниковых формообразований, когда мы, зная время освобождения от ледника тех или иных местностей, можем надежно оценивать период времени, в пределах которого должно было произойти образование подвидов, обитающих сегодня в этих местах (табл. 15.1).
| Таблица 15.1. Примеры скоростей образования новых подвидов | ||
| Вид | Район образования нового подвида | Период, лет |
| Благородный олень (Cervus elaphus) | Великобритания | Около 8 тыс. |
| Ладожская нерпа (Pusa hispida lagopensis) | Оз. Ладога | » 9 тыс. |
| Сайменская нерпа (Pusa hispida saimensi) | Оз. Сайма, Финляндия | » 9 тыс. |
| Хохлатый жаворонок (Galerida cristata) | Низовья р. Нил, Египет | » 8—10 тыс. |
| Сизый голубь (Columba livia) | Оазис Дакла. Северная Африка | » 8—10 |
| Варакушка (Cyanosylvia svecica) | Норвегия | » 8—10 |
| Обыкновенный тюлень (Phoca vitulina) | Северная Канада | » 4 тыс. |
| Европейский кролик (Oryctolagus cuniculus) | О-ва у Великобритании | » 500 |
| Домовая мышь (Mus musculus) | Фарерские о-ва | Не более 3 тыс. |
| 10 подвидов млекопитающих | О. Ньюфаундленд | Не более 12 тыс. |
| 4 подвида рыб | О. Айл-Роял на оз. Верхнее | 12—25 тыс. |
Можно видеть, что подвид часто образуется за несколько тысяч лет, а иногда за сотни или сотни тысяч лет. Полнота палеонтологического материала по отдельным группам в настоящее время настолько велика, что позволяет с уверенностью определять время, необходимое для возникновения новых видов. Особенно удобен для этого анализ последовательных ископаемых форм (см. рис. 6.4; 6.5). Расчеты показывают, что для образования отдельных видов (фратрий) верхнеплиоценовых моллюсков на территории Германии потребовалось несколько миллионов лет, тогда как в Черном море в четвертичном периоде новый вид рода Paphia возник всего примерно за 6000 лет. Один вид зубра (Bison) на Кавказе в среднем плейстоцене существовал примерно 50—60 тыс. поколений, а переход к новому виду занял 10—16 тыс. лет (2—4 тыс. поколений). Оказалось также, что время существования отдельных видов может значительно колебаться — от 0,5 до 5 млн лет, а время существования родов значительно различается, например, у моллюсков и хищных млекопитающих. У моллюсков средняя длительность существования родов — десятки миллионов лет, у хищных — около 8 млн лет. Это означает, что темп формообразования у хищных в десяток раз быстрее, чем у двустворчатых моллюсков. При этом надо учесть, что определяемая таким образом скорость эволюции несколько занижена, так как время видообразования может быть существенно короче, чем время дальнейшего существования вида. Поскольку на палеонтологическом материале надежнее выделяются не виды, а роды и более крупные таксономические единицы, возникает возможность определения скоростей эволюции по числу образованных в единицу времени родов и семейств. На рис. 15.11 приведены такие данные для шестилучевых кораллов на протяжении 550 млн лет — с триаса до неогена. Видно, что в разные геологические периоды скорость эволюции менялась в 3—4 раза: в триасе и юре I семейство кораллов возникало за 5—5,5 млн лет, и за это же время возникало 5—12 родов, то в палеогене — неогена для возникновения семейства требовалось 17—20 млн лет, и на каждое семейство приходилось 40—67 родов. Это позволяет предположить об уменьшении темпов эрогенной эволюции, при увеличении темпов аллогенной эволюции (адаптивной радиации). Конечно, в других группах скорости эволюции могут быть иными.
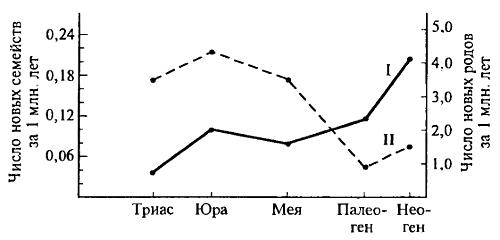
Рис. 15.11. Определение скорости эволюции по числу возникающих новых родов (I) и семейств (II) шестилучевых кораллов Scleractinia (из Д. Раупа и С. Стенли, 1974)
Сравнение большого материала по темпам возникновения и длительности развития новых родов позволило выделить три группы родов (рис. 15.12). К первой группе принадлежат брадителические роды, филогенетические реликты (см. ниже); время их развития превышает 250 млн лет. Ко второй группе принадлежат горотелические роды, время развития которых составляет от нескольких до нескольких десятков миллионов лет, и, наконец, к третьей группе — тахителических родов — принадлежат роды, время развития которых невелико, много меньше I млн лет (Дж. Г. Симпсон).
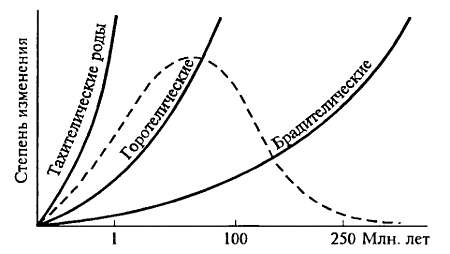
Рис. 15.12. Распределение скоростей эволюции. Три группы родов выделены по темпам возникновения и длительности существования. Большая часть всех изученных родов оказывается горотелическими, а суммарно все роды вместе дают картину, близкую к нормальному распределению (ориг.)
Можно сказать, что ни мутационный процесс, ни волны жизни, видимо, не оказывают решающего влияния на скорость протекания процесса эволюции в любой группе. Но очень велико влияние изоляции и особенно естественного отбора. При определении скорости эволюционного процесса изоляция действует не только как элементарный эволюционный фактор, но и как фактор, активно способствующий перестройке всех биогеоценозов и созданию условий для дальнейшей эволюции. Однако главнейшим фактором, определяющим как продолжительность существования того или иного филума, так и скорость образования новых форм, является естественный отбор.
Успехи молекулярной биологии дали возможность оценить скорость эволюции по замещениям аминокислот в белках и нуклеотидов в нуклеиновых кислотах. Для выражения скорости таких замен принята единица «полинг», равная 10—9 замене на аминокислотный сайт в год (см. гл. 6).
Сравнение аминокислотной последовательности в α- и β-цепях гемоглобина, в молекулах глобулина, цитохрома и других белков показало увеличение числа замен по мере удаления филогенетического расстояния у позвоночных. У всех сравниваемых позвоночных скорость замены для одного и того же белка выражалась сходной величиной в полингах. Разные же белки в пределах позвоночных обычно отличаются по своей скорости эволюции (табл. 15.2). Теперь молекулярно-генетические расчеты позволяют строить филогенетические древа организмов, близкие к морфологическим древам. Скорость молекулярной эволюции (в полингах) не зависит от продолжительности жизни поколения и величины популяций. Постоянство темпов молекулярной эволюции пытаются объяснить фиксацией нейтральных мутаций. Так, на примере α-цепи гемоглобина при сравнении различных позвоночных установлено, что за 7 млн лет происходит замена одной аминокислоты. Данные, полученные по разным белкам, положены в основу идеи молекулярных часов эволюции (см. гл. 6 и 20).
| Таблица 15.2. Скорость эволюции белков у млекопитающих (М. Кимура, 1985) | |
| Белки | Kaa, 10—9 в год |
| Фибринопептиды | 8,3 |
| Панкреатическая рибонуклеаза | 2,1 |
| Лизоцим | 2,0 |
| α-Цепь гемоглобина | 1,2 |
| Миоглобин | 0,89 |
| Инсулин | 0,44 |
| Цитохром C | 0,3 |
| Гистон H4 | 0,01 |
Kaa — среднее число аминокислотных замен, приходящееся на пару гомологичных сайтов двух сравниваемых полипептидов.
