10.2. Предпосылки естественного отбора
Ч. Дарвин обосновал принцип естественного отбора, исходя из двух основных предпосылок: во-первых, характерной для всех живых существ наследственной гетерогенности особей, во-вторых, изначальной, избыточной численности потомства при стационарной общей численности вида в целом.
Гетерогенность особей. Известно, что во всех популяциях особи отличаются друг от друга по многим наследственным особенностям: первично они связаны с возникновением мутаций и комбинированием наследственных факторов родителей при скрещивании. Постоянное действие мутационного процесса и генетической комбинаторики сделало наследственную гетерогенность характерным свойством популяции (см. гл. 7) и первой важнейшей предпосылкой действия естественного отбора.
Прогрессия размножения — другая необходимая предпосылка естественного отбора, обеспечивающая исходную, всегда избыточную численность каждого вида, поскольку на свет появляется гораздо больше особей, чем их доживает до взрослого состояния. На это явление геометрической профессии размножения обращали внимание Ж. Бюффон, К. Линней, Т. Мальтус, А. Бекетов и др., подчеркивая, что каждый вид при реализации его потенциальных возможностей к размножению в состоянии за короткие сроки покрыть всю планету. Однако никто до Ч. Дарвина не понял значения этого явления для эволюции: создания условий для борьбы за существование, с одной стороны, и повышения спектра появляющихся на свет наследственно разнообразных особей — материала для естественного отбора — с другой.
Борьба за существование. Термин «борьба за существование» со времен Ч. Дарвина понимается в метафорическом смысле как любые взаимоотношения особей с окружающими абиотическими и биотическими условиями.
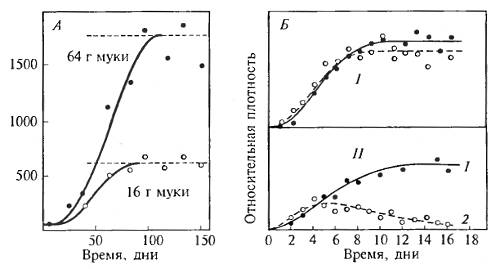
Рис. 10.1. Примеры борьбы за существование (БЗС) в экспериментах. А. Внутривидовая БЗС. Численность мучного хрущака (Tribolium confusum) в зависимости от исходного количества муки (по Р. Чэпман, 1931). Б. Межвидовая БЗС. Численность особей двух видов парамеций (Paramecia) при размножении отдельно (I) и в смешанной культуре (II). В смешанной культуре P.aurelia (1)и P. caudatum (2) второй вид погибает (Г.Ф. Гаузе, 1934)
Борьба за существование охватывает все формы активности особей, направленные на поддержание жизни и размножение. Особь вступает в определенные отношения с другими организмами, фактически соревнуясь в добывании пищи и защите от врагов. Например, заяц спасается от волка и других хищников бегством, но заяц, захвативший зимой при недостатке корма осиновую ветку, соревнуется в этом с другими зайцами. Это примеры проявления прямой борьбы за существование, как межвидовой (зайцы — волки), так и внутривидовой (зайцы — зайцы) (рис. 10.1). Встречается еще косвенная (или соревнование) борьба за существование. Например, особи одного вида как бы соревнуются между собой в устойчивости против действия неблагоприятных факторов среды — голода, холода, засухи, засоленности и т. д. (конституциональная борьба). Примеры борьбы за существование не сводятся к борьбе в прямом смысле слова. Даже внутривидовые отношения в разные периоды и в зависимости от среды могут быть конкурентными, нейтральными или играть роль взаимопомощи. Так, например, при небольшой плотности отношения между личинками дрозофилы нейтральны, при средней плотности они помогают друг другу использовать среду, предотвращая высыхание ее и развитие бактерий и плесени. При очень высоких плотностях отношения между личинками дрозофилы оказываются явно конкурентными в использовании среды. Точно так же в зависимости от освещенности и насыщения почвы элементами питания взаимоотношения между видами бобовых и злаковых переходят от сотрудничества к конкуренции. В экспериментах на примере культурных растений и сорняков показано, что при внесении больших количеств удобрения конкурентные взаимоотношения между ними ослабляются. Обоюдополезные взаимоотношения насекомых и опыляемых ими растений, так же как и взаимоотношения гриба и водоросли в лишайнике, тоже есть формы борьбы за существование.
При более подробной классификации форм борьбы за существование выделяют (И.И. Шмальгаузен и др.) внутригрупповую (которая, в свою очередь, может быть активной и пассивной), межсемейную и межгрупповую (также активную и пассивную). Борьба за существование связана с гибелью (элиминацией) особей. Различают такие типы элиминации: общая, индивидуальная, прямая и косвенная, семейная и групповая. Выделение этих категорий показывает на чрезвычайную сложность проявления борьбы за существование, изучением которой занимаются многие направления современной экологии.
Единственным объективным мерилом борьбы за существование служит соотношение между числом родившихся особей и числом особей из них, принявших участие в оставлении потомства. Но это не что иное, как естественный отбор — дифференциальное размножение особей в популяциях (см. ниже). Поэтому Ч. Дарвин рассматривал борьбу за существование скорее как предпосылку действия естественного отбора.
В эволюционном учении подробный анализ борьбы за существование приобретает смысл только с точки зрения механизма действия естественного отбора. Несомненно, что в рамках экологии специальное изучение роли борьбы за существование, интенсивности ее в разных условиях, а также выяснение связей между организмами в биоценозах может иметь и важное самостоятельное значение. Можно сказать, что в этой области эволюционной экологии приоритет принадлежит отечественным биологам (В.Н. Сукачев, Г.Ф. Гаузе, С.А. Северцов и др.), которые экспериментально в 30-е годы XX в. исследовали роль борьбы за существование.
Концепция борьбы за существование лежит в основе большинства разделов современной экологии и, в частности, представлений о механизмах регуляции численности особей в популяции.
