9.4. Изоляция как элементарный эволюционный фактор
Изоляция — возникновение любых барьеров, ограничивающих панмиксию. Значение изоляции в процессе эволюции и сводится к нарушению свободного скрещивания, что ведет к увеличению и закреплению различий между популяциями и отдельными частями всего населения вида. Без такого закрепления эволюционных различий невозможно никакое формообразование.
Разнообразие форм и проявлений изоляции в природе так велико, что для понимания эволюционной роли изоляции необходимо кратко описать основные ее проявления в природе.
Классификация явлений изоляции. В природе существуют пространственная и биологическая изоляции.
Пространственная изоляция может существовать в разных формах: водные барьеры разделяют население «сухопутных» видов, а барьеры суши изолируют население видов-гидробионтов; возвышенности изолируют равнинные популяции, а равнины — горные популяции и т. д. Сравнительно малоподвижные животные — наземные моллюски на Гавайских островах связаны с влажными долинами. В результате, в каждой из сотен долин на больших островах возникает самостоятельная популяция со своими специфическими особенностями.
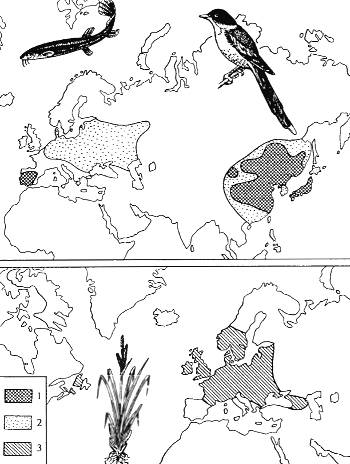
Рис. 9.4. Примеры разорванных ареалов: ареалы голубой сороки (Cyanopica суапа) (1) и вьюна (Misgurnus fossilis) (2) (из Н.В. Тимофеева-Ресовского и др., 1977)
Возникновение территориально-механической изоляции объясняется историей развития видов на определенных территориях. В приведенных двух случаях (рис. 9.4) основной причиной изоляции стало наступление ледников. За время, прошедшее после исчезновения ледников, изолированные формы еще не приобрели значительных морфофизиологических различий и относятся, по-видимому, к единым видам. Впрочем молекулярно-генетические исследования могут показать, что это формы уже возникшего видового ранга (полной изоляции на протяжении жизни десятков тысяч поколений обычно бывает достаточно для возникновения видов).
В настоящее время в связи с деятельностью человека в биосфере все чаще и чаще возникает подобная пространственная изоляция отдельных популяций внутри очень многих видов. Типичным примером стало возникновение в Евразии к началу XX в. разорванного ареала у соболя (Martes zibellina) — результат интенсивного промысла (рис. 9.5). Обычно быстрое возникновение подобного разорванного ареала служит опасным симптомом возможного исчезновения вида.
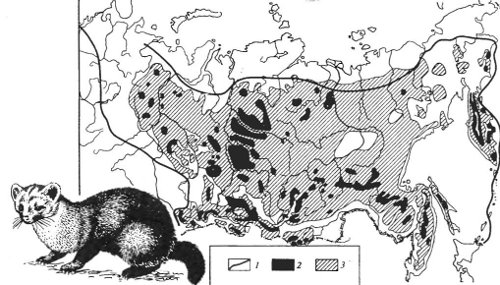
Рис. 9.5. Пространственная структура ареала соболя (Martes zibellina) (по А.А. Насимовичу и В.В. Тимофееву, 1973): 1 — границы ареала; 2 — оставшиеся в 30-е годы популяционные островки населения; 3 — территория, занимаемая соболем в 80-е годы XX
Пространственная изоляция может возникнуть внутри видов малоподвижных животных и растений, не разделенных заметными физико-географическими барьерами. Известно, что обыкновенный соловей (Luscinia luscinia), населяющий многие районы центральной части европейской территории России, в настоящее время практически сплошь находит подходящие условия для гнездования как в не обжитых человеком местах, так и в зарослях по обочинам дорог, в парках и даже скверах больших городов. При этом наблюдается четко выраженная клинальная изменчивость пения этих птиц: по числу «коленец», по тембру и другим особенностям существуют постепенные переходы от одной местности к другой (характер песни наследственно обусловлен). Возникновение такой клинальной изменчивости возможно только потому, что соловьи, несмотря на существование у них частых сезонных перелетов, обладают большим гнездовым консерватизмом: молодые возвращаются практически на то же место, где они вывелись.
Пространственная изоляция внутри вида существует в двух проявлениях: изоляция какими-либо барьерами между частями видового населения и изоляция, определяемая большей возможностью спаривания близко живущих особей, т. е. изоляция расстоянием.
Возникновение пространственной изоляции связано с радиусом репродуктивной активности вида (см. гл. 7).
Радиус репродуктивной активности наземных моллюсков обычно равен нескольким десяткам метров, а чирков — тысяче километров. Значение физико-географических барьеров в пространственной изоляции связано с биологическими особенностями вида в целом.
Биологическую изоляцию обеспечивают две группы механизмов: устраняющие скрещивание (докопуляционные) и изоляция при скрещивании (послекопуляционные). Первые механизмы предотвращают потерю гамет, вторые связаны с потерей гамет и зигот (Э. Майр).
Спариванию близких форм препятствуют различия во время половой активности и созревания половых продуктов. Известно существование «яровых» и «озимых» рас у миног (Lampetra) и некоторых лососевых рыб (Oncorhynchus), которые резко отличаются временем нереста; между особями каждой из рас существует высокая степень изоляции. Среди растений известны случаи генетически обусловленного сдвига в период цветения, создающего биологическую изоляцию этих форм — явление фенологического полиморфизма (рис. 9.6).
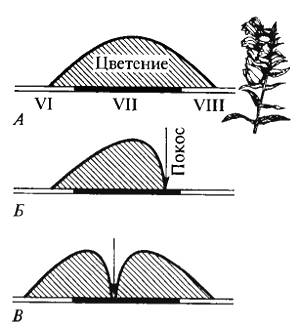
Рис. 9.6. На некошеных лугах большой погремок (Alectorolophus major) цветет и плодоносит в течение всего лета (Л). При регулярном кошении в конце лета (Б) образовалась раса погремка, успевающая принести семена до начала покоса. При более раннем регулярном покосе (В) возникли две расы — ранневесенняя и позднеосенняя. Позднеосенняя раса содержала растения, замедленно развивающиеся до начала покоса, очень низкие, не повреждаемые при косьбе, но затем быстро зацветающие и успевающие дать семена до начала мороза. Пример фенологического полиморфизма (рисунок по данным Н.В. Цингера, 1920)
В природе обычна биотопическая изоляция, при которой потенциальные партнеры по спариванию встречаются, так как они реже предпочитают разные места обитания. Так, часть зябликов (Fringilla coelebs) гнездится в лесах таежного типа, а другая — в невысоких и редких насаждениях с большим числом полян. Потенциальная возможность перекрестного спаривания особей этих групп ограничена. Интересный пример биотопической изоляции — симпатрические внутривидовые формы у обыкновенной кукушки (Cuculus canorus). В Европе обитает несколько «биологических рас» кукушек, различающихся генетически закрепленной окраской яиц. В Восточной Европе одни откладывают голубые яйца в гнезда обыкновенной горихвостки и лугового чекана, другие — светлые в крапинку яйца — в гнезда мелких воробьиных птиц, имеющих яйца сходной окраски. Изоляция между этими формами кукушек поддерживается за счет уничтожения видами-хозяевами недостаточно замаскированных яиц. У многих видов предпочтение биотопа — эффективный изоляционный механизм.
Большое значение в возникновении и поддержании биологической изоляции у близких форм имеет этнологическая изоляция — осложнения спаривания, обусловленные особенностями поведения. Велико разнообразие способов этологической изоляции у животных. Ничтожные на первый взгляд отличия в ритуале ухаживания и обмене зрительными, звуковыми, химическими раздражителями будут препятствовать продолжению ухаживания. Возможно, у животных этологические механизмы — наиболее обширная группа из докопуляционных изолирующих механизмов.

Рис. 9.7. Пример эколого-этологической изоляции. Характеристика световых вспышек североамериканских светляков рода Photurus. Высота и длина пятен в таблице указывают на интенсивность и длительность вспышки (из Э. Майра, 1968)
На рис. 9.7 показан характер световых вспышек у североамериканских светляков рода Photurus. Различия между близкими видами, а иногда и разными группами популяций внутри вида, определяются длительностью, частотой и интенсивностью вспышек.
Важным изолирующим механизмом, затрудняющим скрещивание близких видов, оказывается возникновение морфофизиологических различий в органах размножения (морфофизиологическая изоляция), а также специализация в опылении у растений.
У животных близких видов различия копулятивных органов особенно характерны для некоторых легочных моллюсков, насекомых, а среди млекопитающих — ряда групп грызунов.
Описанные главные механизмы изоляции обычно достаточны для предотвращения в природных условиях возможности скрещивания форм, принадлежащих к разным видам, и понижения эффективности скрещивания у внутривидовых форм, далеко разошедшихся в процессе эволюции (подвидов, групп удаленных географических популяций и т. п.).
Вторая большая группа изолирующих механизмов в природе связана с возникновением изоляции после оплодотворения (собственно-генетическая изоляция), включающей гибель зигот после оплодотворения, развитие полностью или частично стерильных гибридов, а также пониженную жизнеспособность гибридов.
При межвидовом спаривании часто образуются вполне жизнеспособные гибриды, но у них, как правило, не развиваются нормальные половые клетки. В случае же нормального развития гамет гибриды оказываются малоплодовитыми. В природе есть случаи такой изоляции посредством гибридизации: на границе обитания двух близких форм постоянно существует зона, населенная вполне жизнеспособными гибридными особями, но их потомство либо ослаблено и не выдерживает конкуренции с более сильными особями родительских видов, либо нежизнеспособно. На существование такого гибридизационного барьера между видами обычно указывает наличие в природе устойчивых гибридных зон между близкими видами. Такие гибридные зоны известны для некоторых насекомых, серой и черной ворон в Европе (рис. 9.8) и других форм.
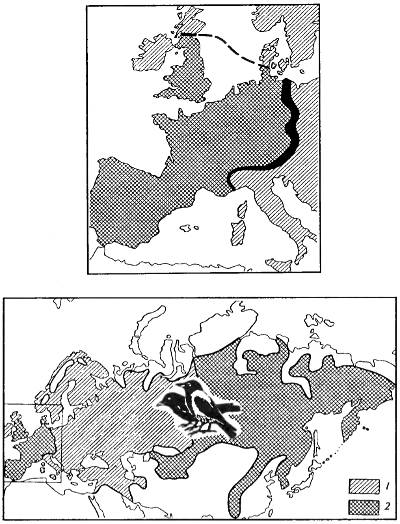
Рис. 9.8. Соприкосновение двух близкородственных форм — черной (Corvus corone) и серой ворон (Corvus corax) без образования широкой гибридной зоны (пример изоляции посредством гибридизации по Э. Майру, 1968). Гибриды оказываются менее жизнеспособными по сравнению с родительскими формами. Судя по неодинаковой ширине гибридной зоны и некоторому колебанию ширины зоны в разные годы, относительная жизнеспособность гибридов, видимо, различна в разных поколениях и частях ареала: 1 — серая ворона; 2 — черная ворона
