8.3. Встречаемость мутаций в природных популяциях
Изучение большого числа природных популяций разных видов, особенно активно развернувшееся в 30—40-е годы XX в., подтвердило вывод С.С. Четверикова (1926) о насыщенности всех популяций разнообразными мутациями. Нет двух популяций, имеющих одинаковые частоты встречаемости и спектры мутантных признаков. При этом близко расположенные, на весьма изолированные, соседние популяции могут отличаться друг от друга столь же значительно, как и далеко расположенные (рис. 8.5).
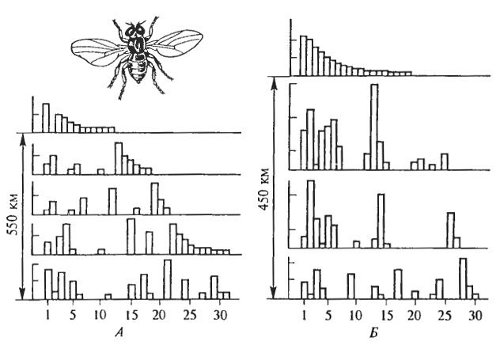
Рис. 8.5. Содержание различных мутаций в географически удаленных (А) и близких (Б) популяциях Drosophila funebris (по Н.В. Тимофееву-Ресовскому и др., 1972)
Особый интерес представляют случаи распространения мутаций в пределах ареала вида, позволяющие судить о том, как мутация «выходит» на историческую, эволюционную арену. На рис. 8.6 приведен один из примеров такого рода: распространение одной полудоминантной мутации слияния пятен на надкрыльях растительноядной божьей коровки, обитающей в Южной Франции, Северной Италии, на Корсике и в Сардинии.
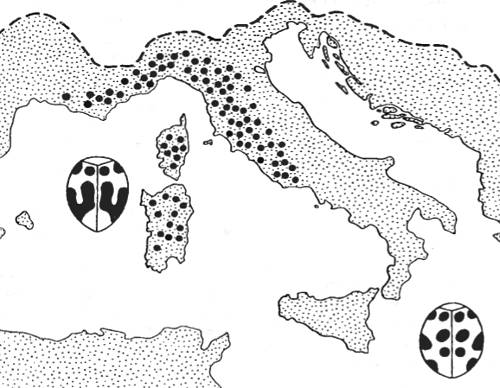
Рис. 8.6. Пример распространения мутации в пределах ареала вида: распространение полудоминантной мутации elaterii (крупными точками), определяющей фен слияния пятен на элитрах божьей коровки (Epilachna chrysomelina) (из Н.В. Тимофеева-Ресовского, 1939)
По изучению генетического состава природных популяций проведено много работ на разных группах растений (львиный зев, фиалка, пикульник и др.), беспозвоночных (дрозофилы, непарный шелкопряд, божьи коровки и др.) и позвоночных (мышевидные хомячки, домовые мыши, крысы, полевки и др.). Во всех случаях популяции отличались друг от друга лишь встречаемостью генных, хромосомных или геномных мутаций. Приведем лишь два характерных примера. На рис. 8.7 приведены данные по встречаемости диагностического признака (второй двойной поперечной кубитальной жилки крыла) у самцов шести видов земляных ос. Эксперименты показали, что этот признак определяется моногибридно наследующейся рецессивной мутацией. У трех видов признак встречается в качестве редкого варианта, у двух других видов — как обычный вариант, наконец, у одного вида — как видовой, встречаясь у большинства особей.
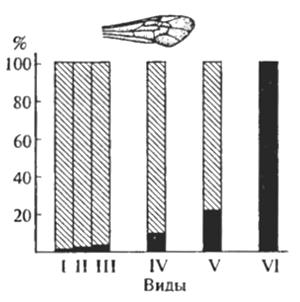
Рис. 8.7. Частота встречаемости второй кубитальной жилки в крыле самцов шести видов земляных ос рода Andrena (по К. Циммерману, 1933)
Другим примером, частично уже рассмотренным в разделе о типах мутаций, может служить распространение разного рода хромосомных мутаций (инверсии) среди популяций у трех видов дрозофилы из Северной Америки (см. рис. 6.27). В результате тщательного цитогенетического анализа и прямых генетических экспериментов удалось восстановить эволюционную последовательность возникновения отдельных групп популяций и видов.
При детальном генетическом сравнении близких природных таксонов (групп популяций, подвидов и близких видов) различия между ними всегда сводились к различиям по наличию отдельных мутаций и их комбинаций. Это дает основание полагать, что и во всех других случаях именно мутации — действительно элементарный эволюционный материал. Особенности разных типов мутаций, достаточно высокая частота их возникновения в природе, изменение ими любых, в том числе и биологически важных признаков, насыщенность природных популяций мутациями свидетельствуют о том, что мутации как элементарные единицы наследственной изменчивости могут рассматриваться в качестве элементарного эволюционного материала.
Этот вывод подтверждается и тем, что именно мутации и их комбинации первично определяют изменение генотипического состава популяции, т. е. возникновение элементарного эволюционного явления.
