ГЛАВА 8
Генетические основы эволюции
Эволюционные изменения признаков и свойств организмов обусловлены изменением генотипов, поэтому понимание основных генетических процессов, происходящих в популяции, необходимо для всей современной эволюционной теории. Успехи генетики позволили классифицировать и изучить основные формы наследственной изменчивости, а также выяснить значение изменчивости разных типов для протекания эволюционного процесса. Изучение роли разнообразных проявлений изменчивости в эволюции позволило сформулировать представление о генетических основах эволюции.
8.1. Изменчивость — свойство органической природы
В основе изменчивости как всеобщего явления живой природы лежит конвариантная редупликация. Именно в процессе матричного копирования ДНК и РНК происходят ошибки последовательности расположения нуклеотидов, связанные с заменой одного нуклеотида другим или сдвигом рамок считывания. Эти процессы лежат в основе изменчивости на молекулярном уровне. Общие причины генетической изменчивости чрезвычайно разнообразны.
Ч. Дарвин писал, что даже в потомстве одной пары родителей особи всегда будут различаться. «Самый опытный натуралист изумился бы многочисленности случаев изменчивости даже самых существенных частей строения». Последующее развитие генетики, раскрыв причины изменчивости, полностью подтвердило эти слова.
Фенотипическая, генотипическая и паратипическая изменчивость. Во времена Ч. Дарвина всю наблюдаемую изменчивость делили на наследственную и ненаследственную. В настоящее время такое разделение правильно лишь в общих чертах. Ненаследственных признаков нет и быть не может: все признаки и свойства организма в той или иной степени наследственно обусловлены. В процессе размножения от поколения к поколению передаются не признаки, а код наследственной информации, определяющий лишь возможность развития будущих признаков в каком-то диапазоне. Наследуется не признак, а норма реакции развивающейся особи на действие внешней среды.
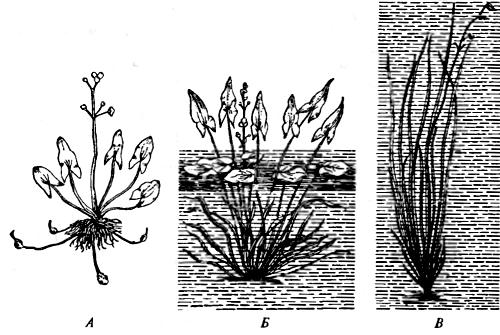
Рис. 8.1. Стрелолист обыкновенный (Sagittaria sagittifolia) формирует разные по форме листья при развитии на суше (А) и в воде (Б). Сигналом для развития водной формы листьев (В) служит не непосредственное действие воды, а затенение (Б. Уоллес, А. Шрб, 1964)
Водный лютик (Ranunculus delphinifolius) и стрелолист (Sagittaria sagittifolia) формируют различные листья под водой и в воздушной среде (рис. 8.1). В свое время Ж. Ламарк использовал этот пример для доказательства изначально адекватного изменения организмов под влиянием условий внешней среды. Однако впоследствии выяснилось, что фактором, определяющим развитие «подводных» листьев, служит не водная среда, а затенение: «пусковым механизмом», определяющим реализацию того или иного варианта в пределах наследственно обусловленной нормы реакций, была закреплена интенсивность света (погруженные в воду листья всегда менее освещены). Поэтому если водный лютик будет развиваться на суше в полутемном помещении, то листья у него будут такими же, какими бывают под водой. Многие растения имеют «световые» и «теневые» листья. У рачка Artemia salina с изменением солености воды меняется число члеников брюшка.
Во всех подобных случаях возможные пределы изменения не случайны, а определяются генотипом, его индивидуальной наследственной программой развития — нормой реакции.
Вся наблюдаемая изменчивость какого-либо признака или свойства в пределах нормы реакции называется фенотипической. Фенотип — совокупность всех внутренних и внешних структур и функций данной особи, развивающаяся как один из возможных вариантов реализации нормы реакции в определенных условиях. В общей фенотипической изменчивости популяции могут быть выделены две доли: генотипическая, или наследственная, и паратипическая, вызванная внешними условиями. Доля общей изменчивости, которая определяется генотипическими различиями между особями по данному признаку, характеризует наследуемость этого признака.
Известно, что жирность молока, содержание белка в молоке и общая молочная продуктивность особей в стаде рогатого скота (Bos taurus) обнаруживают высокую фенотипическую изменчивость. Величина наследуемости для первых двух признаков высока и колеблется от 60 до 70%, тогда как наследуемость общей продуктивности (величина удоя) не превышает 33%. Соответственно этому эффективность отбора оказалась различной для указанных признаков: путем отбора удается сравнительно быстро добиться увеличения жирности и содержания белка в молоке, а отбор на величину удоя малоэффективен (величина удоя зависит в основном от условий содержания животных). Аналогичные результаты наблюдаются при анализе признаков продуктивности кур (Gallus domestica): яйценоскости и массы яйца. У большинства пород (популяции) кур генотипическая составляющая (наследуемость) яйценоскости невелика (12—30%), а наследуемость массы яйца значительна (60—74%). Поэтому отбор в направлении увеличения яйценоскости обычно неэффективен, тогда как отбор на повышенную массу яйца сразу же дает положительные результаты. Однако японские селекционеры обнаружили популяцию кур, для которой показатель генотипической составляющей яйценоскости оказался заметно увеличенным. В результате умелого отбора из этой популяции сейчас выведена порода кур, дающих более 500 яиц в год.
В приведенных примерах высокая доля генотипической изменчивости популяции определила успех селекции. В природных условиях доля генотипической и паратипической составляющих в общем спектре фенотипической изменчивости для большинства видов обычно неизвестна. Однако по аналогии с генетически изученными видами (или генетически проанализированными отдельными признаками и свойствами близких видов) можно с той или иной степенью достоверности предполагать характер наследуемости основных групп признаков.
Внутрипопуляционная изменчивость слагается из разнообразных выражений нормы реакции по любому признаку или свойству. Изменения наследственного материала — мутации — представляют собой элементарный эволюционный материал.
