2.7.3. Семенные растения
Самая процветающая группа растений образует семена. Эти растения, по-видимому, произошли от вымерших семенных представителей папоротников и их близких родичей. Систематика и основные признаки семенных растений суммированы в табл. 2.10.
Таблица 2.10. Систематика и основные признаки семенных растений
| Семенные растения | |
| Основные признаки Спорофит является доминирующим поколением; гаметофит крайне редуцирован Спорофит разноспоровый, т.е. образует споры двух типов: микроспоры и мегаспоры; микроспора – пыльцевое зерно, мегаспора – зародышевый мешок Зародышевый мешок (мегаспора) остается целиком закрытым в семязачатке (мегаспорангии); оплодотворенный семязачаток представляет собой семя* Вода для полового размножения не нужна, поскольку мужские гаметы не способны плавать (исключение составляют некоторые примитивные представители); чтобы оплодотворить яйцеклетки, они проникают в завязь через пыльцевую трубку* Сложные по строению проводящие ткани имеются в корнях, стеблях и листьях |
|
| Отдел Coniferophyta (хвойные) | Отдел Angiospermophyta (цветковые растения) |
| Обычно образуют шишки, на которых развиваются спорангии, споры и семена Семена не спрятаны в завязи. Они лежат на поверхности специализированных листьев, называемых семенными чешуями*, которые собраны в шишки Не образуют плодов, поскольку не имеют завязи |
Образуют цветки, в которых развиваются спорангии, споры и семена* Семена спрятаны в завязи* После оплодотворения из завязи развиваются плоды* Классы: двудольные и однодольные (см. табл. 2.11) |
* Диагностический признак
В табл. 2.10 рассматриваются две основные группы семенных растений – голосеменные и покрытосеменные. Последние чаше называют цветковыми растениями. У голосеменных семязачатки (и впоследствии семена) располагаются на поверхности особых чешуйчатых листьев, называемых мегаспорофиллами или семенными чешуями. Эти чешуи собраны в шишки. У покрытосеменных семязачатки, а следовательно, и семена, заключены в особые структуры, т.е. лучше защищены.
2.7.4. Отдел Coniferophyta (хвойные)
Основные признаки Coniferophyta суммированы в табл. 2.10.
Хвойные (голосеменные) – процветающая группа растений, распространенных по всему земному шару; на их долю приходится примерно одна треть всех лесов планеты. Это деревья и кустарники, в основном вечнозеленые с игловидными листьями. Подавляющее число видов обитают в высоких широтах и распространяются на север дальше всех других деревьев. Хвойные имеют большое хозяйственное значение в качестве «мягкой древесины», используемой не только для получения лесо- и пиломатериалов, но и для получения смолы, скипидара и древесной мезги. К хвойным относятся сосны, лиственницы (с опадающей на зиму листвой), пихты, ели и кедры. Типичный представитель хвойных – сосна обыкновенная (Pinus sylvestris).
Этот вид распространен на всей территории центральной и северной Европы, бывшего СССР и Северной Америки. Он был интродуцирован и в Великобританию, но в природных условиях растет только в Шотландии. Это красивое величественное дерево высотой до 36 м с характерной отслаивающейся корой розовато-или желтовато-коричневого цвета выращивают и для декоративных целей, и для получения пило- и лесоматериалов. Сосны чаще всего растут на песчаных или бедных горных почвах, и поэтому корневая система у них обычно расстилается в поверхностных слоях почвы и сильно ветвится. Внешний вид сосны показан на рис. 2.39.
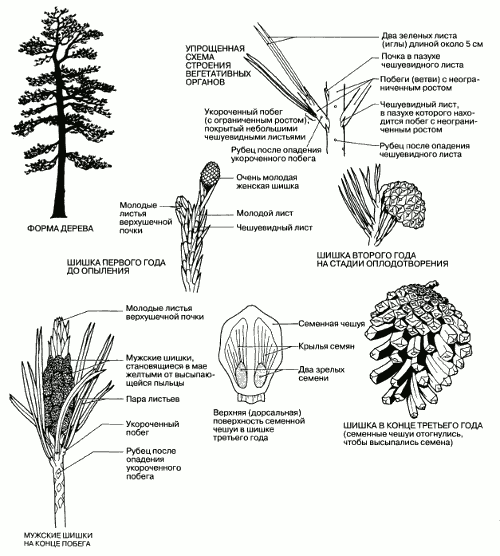
Рис. 2.39. Внешний вид и характерные признаки спорофитного поколения сосны обыкновенной (Pinus sylvestris).
Каждый год из мутовки боковых почек на верхушке ствола вырастает новая мутовка ветвей. Характерный заостренный на конус облик сосны и других хвойных обусловлен тем, что мутовки более коротких (более молодых) веток на верхушке книзу постепенно сменяются все более длинными (более старыми). С возрастом нижние ветви отмирают и отпадают; поэтому крона у старых деревьев обычно сохраняется только на верхушке (рис. 2.39).
Главные ветви и ствол продолжают расти из года в год в результате роста верхушечной почки. Поэтому говорят, что для хвойных характерен неограниченный рост. Чешуевидные листья расположены спирально; в пазухах таких листьев находятся почки, из которых развиваются короткие веточки (длиной 2–3 мм), называемые укороченными побегами. Это – стебли с ограниченным ростом, на верхушке которых растет по два листа. Как только побег вырастает, чешуевидный лист у основания отпадает и на его месте остается рубец. Листья похожи на иголки, что уменьшает площадь их поверхности, а следовательно, и потери воды. Кроме того, листья покрыты толстой восковой кутикулой, а устьица погружены глубоко в ткань листа – еще одна адаптация для сохранения воды. Эти ксероморфные черты позволяют сводить к минимуму потери воды из вечнозеленых листьев во время холодных сезонов, когда вода может замерзнуть, или когда ее трудно извлечь из почвы. Через два-три года укороченные побеги отпадают вместе с листьями и на их месте остается еще один рубец.
Дерево представляет собой спорофитное поколение. Весной на одном и том же дереве образуются мужские и женские шишки. Диаметр мужских шишек составляет около 0,5 см; они округлые и располагаются группами у основания новых побегов под верхушечной почкой. Развиваются они в пазухах чешуевидных листьев вместо укороченных побегов. Женские шишки появляются в пазухах чешуевидных листьев на конце новых сильных побегов на некотором расстоянии от мужских шишек и располагаются более беспорядочно. Поскольку полное развитие шишки занимает три года, шишки очень различаются по размерам, и на одном дереве можно обнаружить шишки от 0,5 до 6 см величиной. Молодые шишки имеют зеленый цвет, но на второй год они становятся коричневыми или красновато-коричневыми. И мужские, и женские шишки состоят из плотно прижатых друг к другу спорофиллов (модифицированных листьев), расположенных спирально вокруг центральной оси (рис. 2.39).
На нижней поверхности каждого спорофилла мужской шишки находятся два микроспорангия, или пыльцевых мешка. Внутри каждого пыльцевого мешка происходит мейотическое деление, приводящее к образованию гаплоидных пыльцевых зерен, или микроспор. В них находятся мужские гаметы. У пыльцевых зерен имеется по два больших воздушных мешка, облегчающих их распространение ветром. В мае шишки становятся совсем желтыми из-за пыльцы, которая целым облаком вылетает из них. В конце лета они увядают и отпадают.
Каждый спорофилл женской шишки состоит из нижней кроющей чешуйки и более крупной верхней чешуи, несущей семязачатки. На верхней поверхности крупной чешуи бок о бок расположены два семязачатка, внутри которых происходит образование женских гамет. Опыление происходит уже на первом году развития шишки, но оплодотворение задерживается до следующей весны, когда вырастут пыльцевые трубки. Из оплодотворенных семязачатков образуются крылатые семена. Созревание семян продолжается в течение второго года, и лишь на третий год они высыпаются. К этому времени шишка становится относительно крупной и одревесневает, а чешуи отгибаются наружу, обнажая семена перед тем как их развеет ветром.
2.7.5. Отдел Angiospermophytae (покрытосеменные, или цветковые растения)
Основные признаки покрытосеменных перечислены в табл. 2.10.
Таблица 2.10. Систематика и основные признаки семенных растений
| Семенные растения | |
| Основные признаки Спорофит является доминирующим поколением; гаметофит крайне редуцирован Спорофит разноспоровый, т.е. образует споры двух типов: микроспоры и мегаспоры; микроспора – пыльцевое зерно, мегаспора – зародышевый мешок Зародышевый мешок (мегаспора) остается целиком закрытым в семязачатке (мегаспорангии); оплодотворенный семязачаток представляет собой семя* Вода для полового размножения не нужна, поскольку мужские гаметы не способны плавать (исключение составляют некоторые примитивные представители); чтобы оплодотворить яйцеклетки, они проникают в завязь через пыльцевую трубку* Сложные по строению проводящие ткани имеются в корнях, стеблях и листьях |
|
| Отдел Coniferophyta (хвойные) | Отдел Angiospermophyta (цветковые растения) |
| Обычно образуют шишки, на которых развиваются спорангии, споры и семена Семена не спрятаны в завязи. Они лежат на поверхности специализированных листьев, называемых семенными чешуями*, которые собраны в шишки Не образуют плодов, поскольку не имеют завязи |
Образуют цветки, в которых развиваются спорангии, споры и семена* Семена спрятаны в завязи* После оплодотворения из завязи развиваются плоды* Классы: двудольные и однодольные (см. табл. 2.11) |
* Диагностический признак
Покрытосеменные лучше других растений адаптированы к жизни на суше. Появившись в меловом периоде примерно 135 млн. лет назад, они быстро вытеснили хвойные, заняв главенствующее положение среди растительности планеты и освоив самые разнообразные местообитания. Некоторые из покрытосеменных вернулись к пресноводному, а несколько видов – солоноватоводному образу жизни.
Одна из самых характерных особенностей покрытосеменных, помимо закрытых семян, о которых уже говорилось, это – появление цветков вместо шишек. Наличие цветков позволило этим растениям привлечь для опыления насекомых, а иногда даже птиц и летучих мышей. Яркая окраска цветков, аромат, съедобная пыльца и нектар – все это средства для привлечения животных-опылителей. В некоторых случаях эволюция опылителей и цветков шла параллельно, что привело к возникновению многих высокоспециализированных и взаимовыгодных отношений. Адаптации цветка, как правило, были направлены на максимальное увеличение шансов для переноса пыльцы насекомыми, и поэтому этот процесс более надежен, чем опыление ветром. Растениям, опыляемым насекомыми, не нужны такие большие количества пыльцы, как при опылении ветром. Тем не менее многие цветковые растения приспособились к опылению ветром.
Двудольные и однодольные
Покрытосеменные растения делятся на две большие группы, которым можно дать статус классов.
Таблица 2.11. Основные различия между двудольными и однодольными
| Класс Dicotyledoneae | Класс Monocotyledoneae | |
| Примеры | Горох, роза, лютик, одуванчик | Злаки, ирис, орхидеи, лилии |
| Морфология листа | Сетевидный рисунок жилок (сетчатое жилкование) | Жилки параллельны (параллельное жилкование) |
| Лист дифференцирован на листовую пластинку и черешок | Обычно длинные и тонкие подобные листьям злаков (рис. 2.40) | |
| Дорсальная и вентральная поверхности различаются | Дорсальная и ветральная поверхности одинаковы | |
| Анатомия стебля | Кольцо из проводящих пучков | Проводящие пучки разбросаны |
| В проводящих пучках обычно имеется камбий, обеспечивающий вторичный рост | Камбия в проводящих пучках обычно нет, поэтому вторичный рост не наблюдается (имеются исключения, например пальмы) | |
| Морфология корня | Первичный корень (первый корень, выросший из семени) сохраняется в виде центрального стержневого корня, от которого отходят боковые корни (вторичные корни) | Придаточные корни, отходящие от основания стебля, не отличаются от первичного корня; корневая система мочковатая |
| Анатомия корня | Несколько групп ксилемы (2–8)(см. гл. 13) | Множество групп ксилемы (обычно до 30) |
| Часто в проводящих пучках имеется камбий, обеспечивающий вторичный рост | Камбия в проводящих пучках обычно нет, поэтому не происходит и вторичного роста | |
| Морфология семени | Зародыш имеет две семядоли (листья, развивающиеся в семени) | У зародыша одна семядоля |
| Цветки | Число частей в основном кратно 4 и 5 | Число частей кратно 3 |
| Чашечка и венчик обычно различаются | Нет явного деления на чашечку и венчик. Эти структуры часто соединены, образуя «сегменты околоцветника» | |
| Часто опыляются насекомыми | Часто опыляются ветром |

Рис. 2.40. Строение листа однодольных (А) и двудольных (Б).
Чаще всего эти две группы называют однодольными и двудольными. В табл. 2.11 перечислены основные признаки, по которым они различаются (см. также рис. 2.40). Согласно современным представлениям, однодольные, вероятно, произошли от двудольных.
Покрытосеменные бывают травянистые (т.е. не одревесневшие) и деревянистые. К последним относятся кустарники и деревья. У этих растений образуется большое количество вторичной ксилемы (древесины), которая выполняет опорную функцию и, кроме того, служит проводящей тканью. Ксилема возникает в результате активности проводящих пучков камбия. Этот камбий представляет собой слой клеток, расположенных между ксилемой и флоэмой в стеблях и корнях. Клетки камбия сохраняют способность к делению. Образующаяся при этом новая ксилема называется вторичной ксилемой или древесиной.
Травянистые растения, или травы, полагаются для опоры только на тургесцентность клеток и на большое количество механических тканей, таких как колленхима, склеренхима и ксилема; неудивительно поэтому, что и сами они не очень велики. У травянистых растений либо совсем нет камбия, либо, если он и имеется, его активность незначительна.
Многие травянистые растения однолетние, т.е. весь цикл развития от семени до семени у них завершается за один год. У некоторых травянистых растений образуются многолетние органы типа луковиц, клубнелуковиц или клубней, благодаря которым растение перезимовывает или же переживает периоды неблагоприятных условий, например засуху (гл. 20). Такие травянистые растения могут быть двулетними или многолетними. В первом случае они и образуют семена, и отмирают на второй год, во втором – живут много лет. Кустарники и деревья – растения многолетние. Они могут быть либо вечнозелеными, либо листопадными. Вечнозеленые растения образуют и сбрасывают листву круглый год, поэтому на растениях всегда есть листья. Листопадные полностью сбрасывают листья в холодное или засушливое время года.
Разнообразие покрытосеменных проиллюстрировано на рис. 2.41–2.44 на примере типичных представителей этой группы растений.
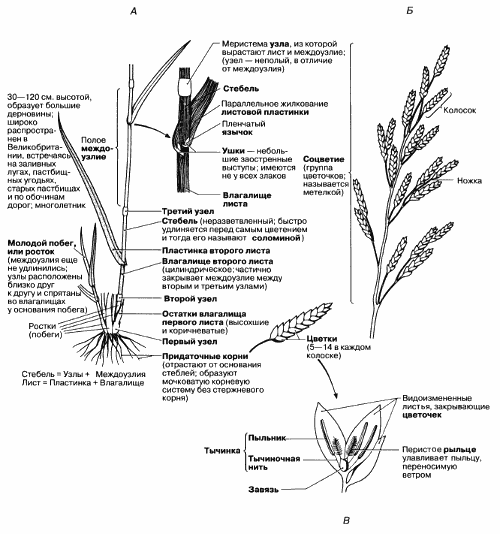
Рис. 2.41. Строение цветка и вегетативных органов однодольного травянистого растения овсяницы луговой (Festuca pratensis). Вторые листья обозначены на рисунке серым цветом. Листья обычно расположены двумя рядами попеременно то на одной, то на противоположной стороне стебля. А. Строение вегетативных органов. Б. Строение соцветия. В. Детали строения одиночного открытого цветка, или цветочка; пленочки, или лодикулы, – две небольшие лепестковидные структуры, покрывающие завязь, на рисунке не показаны.
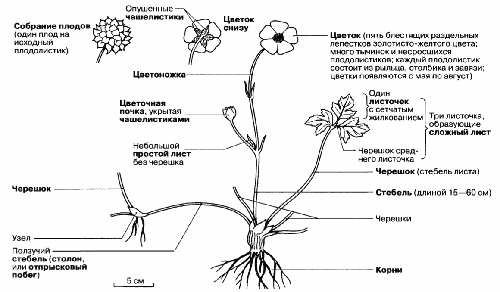
Рис. 2.42. Строение цветка и вегетативных органов двудольного травянистого растения лютика ползучего (Ranunculus repens). Это обычное многолетнее растение встречается на мокрых лугах, в сырых лесах, садах и на брошенных пастбищах по всей Великобритании.
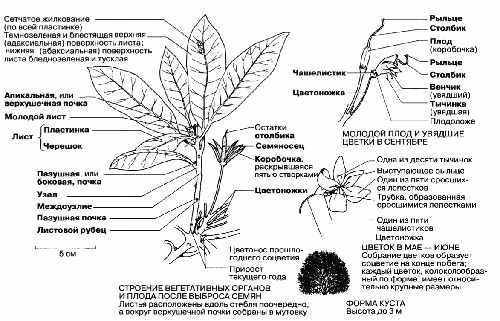
Рис. 2.43. Строение цветка и вегетативных органов дикого рододендрона (Rhododendron ponticum) – вечнозеленого двудольного кустарника. Исходно интродуцированное, это растение успешно акклиматизировалось и хорошо растет на кислых почвах (песчаных и торфяных), на пустошах и в лесах.
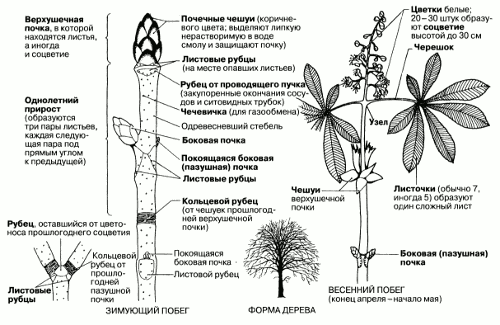
Рис. 2.44. Строение цветка и вегетативных органов конского каштана (Aesculus hippocastanum), двудольного листопадного дерева, достигающего в высоту 30 м и более.
