Иммунологические методы. Биохимия располагает специальными методами исследования, позволяющими с большой степенью точности выяснить «кровное родство» разных групп. При сравнении белков крови используется способность организмов вырабатывать (в ответ на введение в кровь чужих белков) антитела, которые затем можно выделить из сыворотки крови и определить, при каком разведении эта сыворотка будет реагировать с другой, сравниваемой сывороткой (реакции преципитации). Такой анализ показал, например, что ближайшие родственники человека – высшие человекообразные обезьяны, а наиболее далекие среди приматов – лемуры. Обнаружено серологическое сходство крови хищных млекопитающих с ластоногими, близость птиц к крокодилам и черепахам (птицы оказываются филогенетически ближе к крокодилам, чем крокодилы к змеям). Показаны глубокие серологические различия между хвостатыми и бесхвостыми амфибиями; найдены черты сходства по реакции преципитации между хордовыми и иглокожими и т.д. Подобные методы исследований применимы и к растениям.
Биохимический полиморфизм. Начиная с 60-x годов в практику микроэволюционных исследований широко входит метод изучения вариаций белков, обнаруживаемых с помощью электрофореза (Р. Левонтин). С помощью этого метода можно с большей точностью, чем прежде, определять уровень генетической изменчивости в популяциях, а также степень сходства и различия между популяциями по сумме таких вариаций.
Различия между популяциями по биохимическим показателям можно наглядно определять на карте (рис. 6.30) либо на дендрограмме, построенной по коэффициентам сходства и различий (рис. 6.31).
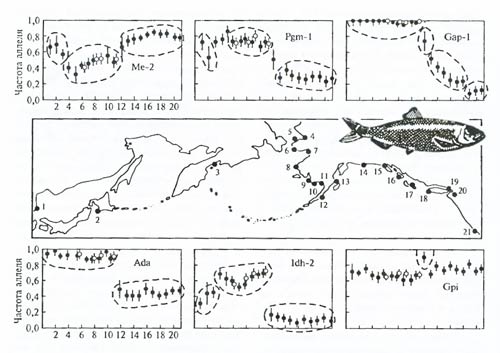
Рис. 6.30. Пример феногеографического анализа по частотам аллозимов (Ме-2, Pgm-l,Gap-l, Ada, Idh-2, Gpi) популяционной структуры тихоокеанской сельди (Clupea pallasi) (по В. Грант, Ф. Уттер, 1984).
1–21 – места сбора материала, пунктиром выделены возможные популяционные группировки
По частотам электроморф можно исследовать внутрипопуляционную пространственно-генетическую структуру и выявлять динамику этой структуры, т.е. исследовать начальные стадии процесса микроэволюции (см. гл. 8, 9).
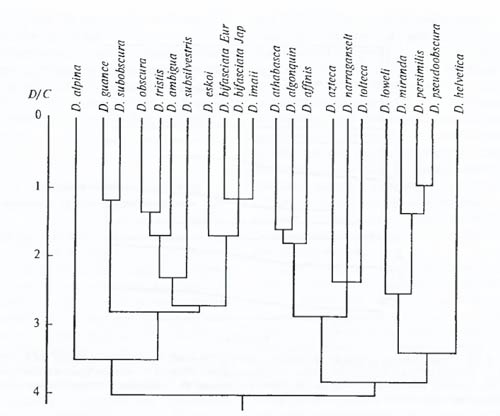
Рис. 6.31. Пример построения филогенетических отношений в виде дендрограммы на основании данных электрофореза белков: взаимоотношения между видами дрозофил группы D. pseudoobscura. D/C – среднее число аминокислот, по которым различаются ферменты разных видов (из С.М. Гершензона, 1979)
При биохимических сравнениях ныне живущих организмов показано, что ранние метаболические звенья синтеза стеринов, каротиноидов, вторичных метаболитов, реакции гликолиза и дыхания у многих растений и животных сходны и не нуждаются в участии кислорода. Завершающие же стадии многих метаболических путей, особенно у высокоорганизованных существ, протекают с его участием. Это свидетельствует об их эволюционной молодости и сложении в аэробной фазе истории Земли. Таким образом, с помощью биохимических методов (и в том числе молекулярно-биологических) можно исследовать эволюционные процессы самого разного масштаба. Несомненно, что в ближайшие годы арсенал этих методов должен существенно расшириться, учитывая значительное развитие исследований в этой области.
Методы моделирования эволюции. В последние десятилетия появилась возможность моделировать эволюционный процесс. Конечно, сложность организации жизни такова, что пока невозможно надежное моделирование развития экосистемы или целой группы, но влияние отдельных эволюционных факторов на популяцию и вид в целом поддается моделированию. Пока такое моделирование неизбежно связано с принятием целого ряда упрощающих условий (например, рассматриваются обычно лишь двулокусные генетические системы, принимается равновероятной возможность скрещивания особей в популяции друг с другом и другие упрощения, далекие от природных ситуаций). Но и при таких ограничениях моделирование иногда помогает уже сейчас решать ряд эволюционных задач. Моделирование эволюционного процесса позволяет, в частности, исследовать возможные влияния отдельных факторов на ход эволюции, их взаимодействие, выяснить в некоторых случаях возможные направления и результаты процесса эволюции (рис. 6.32). Математическое моделирование показало (А.Н. Колмогоров, 1935), что наибольшие темпы изменения популяций должны быть не в больших и не в малых изолированных популяциях, а в средних по величине, с периодически возникающими потоками генов между ними.
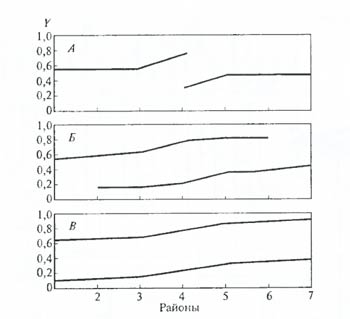
Рис. 6.32. Пример моделирования эволюции: распространение признака Y (форма рыла: острорылость – тупорылость) в популяциях ленка Brachymysrax в Азии: А – стадия расхождения признаков в районе первого контакта; Б – промежуточное состояние; В – возникшее стационарное состояние по данной паре признаков (по М.В. Мины, 1985)
При применении быстродействующих ЭВМ с большой памятью возникает возможность как бы резко ускорить «процесс эволюции» и составить прогноз развития эволюционных событий при различном сочетании условий (см. также раздел 15.8).
Особое внимание при моделировании процесса эволюции приходится уделять выбору необходимых и достаточных исходных предпосылок. Можно заложить в машину и «проиграть» различнейшие варианты взаимодействия тех или иных эволюционных сил в той или иной «обстановке», но если упустить какой-либо один из ведущих компонентов, то результаты моделирования могут получиться обратные реально существующим в природе. Определить набор необходимых и достаточных предпосылок для создания модели – дело очень сложное. Такие работы целесообразно организовывать только в тесном контакте математиков и биологов.
Другие методы изучения эволюции. На основании данных, полученных сравнительной физиологией, можно проследить эволюционное развитие тех или иных функций. Так, по обмену веществ грибы ближе стоят к животным, чем к растениям (А.Н. Белозерский), что служит еще одним основанием для выделения их в самостоятельное царство.
Значительный интерес для выяснения тонких особенностей протекания процесса эволюции представляет изучение поведения животных (этология), на что много внимания обращал Ч. Дарвин. У близких видов, недавно дивергировавших от общего предка, сохраняется (или рекапитулирует в процессе онтогенеза) множество общих мелких черт поведения (К. Лоренц, Н. Тинберген и др.). Некоторые характерные древние черты поведения у птиц и млекопитающих могут быть не менее важными показателями происхождения от общих предков, чем гомологичные органы или другие классические сравнительно-анатомические доказательства.
В некоторых случаях эффективным оказывается использование паразитологического метода изучения эволюции. Многочисленными исследованиями доказано, что эволюция паразитов и хозяев протекает сопряженно. Известно также, что в некоторых группах паразиты оказываются специфическими для видов, родов или семейств. Поэтому по присутствию определенных паразитов порой можно с большой точностью судить о филогенетических связях видов – хозяев этих паразитов.
По-видимому, любая биологическая дисциплина обладает своим специфическим набором методов изучения эволюционного процесса, и достаточно полное их перечисление практически невозможно в рамках одного курса.
