Экосистемная теория эволюции (ЭТЭ)
Интуитивно к построению такой теории ученые стремились с конца XIX в. Факты смены биот, зарегистрированные палеонтологической летописью, следы геологических катаклизмов, происходивших на нашей планете на протяжении ее истории, всегда побуждали эволюционно мысливших исследователей к поиску между этими рядами событий причинной связи. В знакомых нам гипотезах сопряженной эволюции была выявлена одна из фундаментальных особенностей биологической эволюции – ее периодичность, или этапность. В разных гипотезах, как мы видели, главные события в истории органического мира связывались с разными внешними причинами – от процессов горообразования до изменений в уровне солнечной и космической радиации.
Наиболее известную экосистемную гипотезу эволюции разрабатывает (с 1969 г.) в России палеобиолог В.А. Красилов. Это целостная концепция эволюции, построенная с последовательно системных позиций и опирающаяся на данные современной молекулярной биологии и генетики. Но это, как отмечает сам автор, всего лишь гипотеза, поскольку единой схемы экосистемной эволюции пока не существует. Остановимся на ней подробнее.
Движущей силой изменения и эволюции биосферы выступает ее взаимодействие с геологическими оболочками Земли, периодически принимающее характер кризиса. Конечно, эволюционные преобразования в биосфере совершаются и в спокойные межкризисные периоды, но их масштабы неизмеримо скромнее. Для быстрых и радикальных перемен необходим толчок извне. Когда он возникает и оказывается воспринятым биосферой, импульсы от нее по нисходящей каузальной цепочке идут к подчиненным ей системам – биоценозам, популяциям, генофондам. Наиболее вероятен следующий порядок развертывания геологических событий (рис. 29).
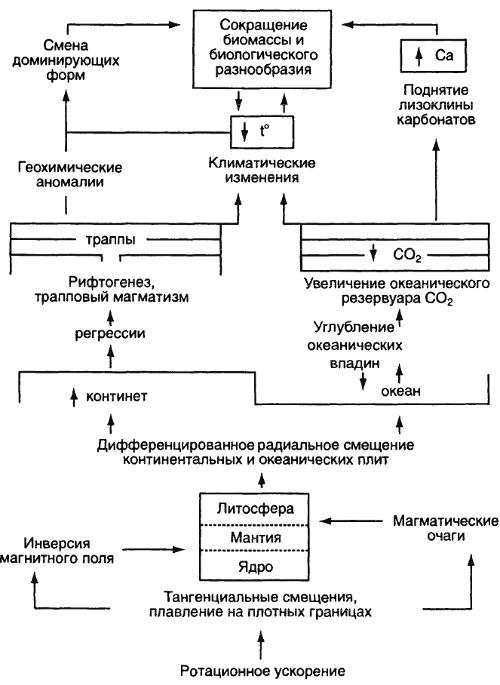
Рис. 29. Схема развития кризисных событий (из: Красилов, 2001)
В силу постоянно меняющегося взаиморасположения небесных тел Земля испытывает периодические колебания параметров орбиты, положения оси вращения и угловой скорости вращения. Благодаря инерции ротационные силы Земли порождали растрескивание земной коры, происходившие особенно легко в зоне океанов, где она была более тонкой. По образовавшимся трещинам начинались сдвиги крупных литосферных блоков, которые порождали мощные тектонические процессы, вулканизм, излияние магмы, морские трансгрессии и регрессии и изменения климата. Важно иметь в виду, что изменения ротационного режима нашей планеты в той или иной степени затрагивали все компоненты биосферы и все подчиненные ей экосистемы. Принимая такую последовательность событий, Красилов опирался не только на последние достижения геофизики, но и на гипотезу палеонтолога Личкова (1965), который, как мы знаем, первым обосновал идею об инициирующем воздействии периодических колебаний скорости вращения Земли на крупномасштабные геолого-географические события. Напомним, что он же увязал эти события с хронологически совпадающими с ними революционными преобразованиями биоты.
Среди упомянутых звеньев геологической цепочки для эволюционной судьбы биосферы наиболее значимо последнее – циклические изменения климата, а также тесно связанные с ними процессы горообразования. О роли последних Красилов пишет так: «Приуроченность эволюционных рубежей к тектоническим и климатическим перестройкам отражает основную закономерность эволюционного процесса… Эти перестройки не только влияли на ход эволюции (в чем все так или иначе согласны), но были ее основной движущей силой» (Красилов, 1977. С. 171). Только благодаря им возможно само «естественное членение геологической истории, причем ранг стратиграфических подразделений, очевидно, отвечает масштабу климатических циклов» (Красилов, 1973. С. 238).
Климатические циклы имеют, однако, и другой источник возникновения. Это ритмы солнечной активности, в особенности крупномасштабные циклы в 30 и 180 млн лет. Они фактически совпадают с периодичностью геологических дистроф, и в их существовании теперь мало кто сомневается. Солнечная радиация – мощнейший фактор тотального воздействия на все земные процессы, и колебания ее активности, наряду с изменениями климата, способны породить в биоценозах стрессовую ситуацию.
Кроме того, изменения солнечной активности – главная причина геомагнитных бурь, влияющих на динамику магнитного поля Земли. К изменениям же этого параметра все живое особенно чувствительно. Ныне достоверно установлено, что магнитное поле становилось переменным (меняло свою полярность) как раз в кризисные периоды на рубежах соответствующих эр и периодов. Как нам представляется, Красилов несколько недооценивает значение солнечных ритмов, уделяя им мало внимания.
Геологический кризис, в отличие от процессов, совершающихся в биоценозах, развивается медленно и долгое время носит колебательный характер. За миллионы лет до того, как выйти на уровень биосферы, он зарождается где-то глубоко в недрах Земли, чтобы, окрепнув и развившись, воздвигнуть на Земле горные цепи и произвести все дальнейшие перемены.
Обратим внимание, что в современной ЭТЭ изменения климата и всех прочих физических параметров среды не выступают больше непосредственной причиной преобразований биоты, как это было в старых гипотезах первой половины XX в. Они рассматриваются теперь не более как пусковой механизм, дающий старт дестабилизационным процессам в экосистемах, к рассмотрению которых мы и переходим.
Все познается в сравнении. Чтобы понять, что происходит в биоценозах в условиях кризиса, необходимо иметь представление об их стабильном состоянии. Этому помогло сопоставление тенденций эволюционного развития биоценозов, наблюдаемых в тропиках и в средних широтах с их выраженной сменой сезонов. Как известно, по мере продвижения к экватору экологическая емкость (плотность заполнения видами) биоценозов возрастает, а при движении в обратном направлении – падает. Из этого факта Красилов делает вывод, что при похолодании «биоценозы оказываются перенасыщенными, а при потеплении – недонасыщенными» (Красилов, 1977. С. 236). В первом случае ввиду элиминации избыточных видов и преимущественного распространения видов, способных обеспечивать быстрый рост популяций, структура биоценозов испытывала упрощение, а шансы таксонов на перестройки мегаэволюционного плана снижались. Во втором случае за счет размножения видов, отличающихся более эффективным использованием наличных ресурсов, дробления экологических ниш и прогрессирующей специализации шло усложнение биоценозов и становление новых типов организации (типогенез, или возникновение анастроф).
В спокойные периоды эволюция экосистем происходит медленно и постепенно. Их самая характерная черта – высокая стабильность, присущая всем климаксовым сообществам, завершившим экологическую сукцессию. Стабильность обеспечивают в первую очередь доминантные виды с длительными жизненными циклами и малым числом потомков. В таких сообществах развиты разнообразные механизмы ослабления конкуренции, в полной мере действует стабилизирующий отбор, высок уровень генетической разнородности популяций.
В экосистемах, не завершивших своего развития, благодаря продолжающемуся росту видового разнообразия, удлинению пищевых цепей, углублению специализации и т.п. происходит усложнение структуры. Но эти процессы небезграничны. Им кладут предел ограниченность энергетических ресурсов (Гиляров, 1973) и критическая величина популяции, уменьшение которой грозит виду вымиранием. В насыщенных сообществах эволюция теоретически прекращается.
Палеонтологическая летопись дает достаточно примеров подобных зрелых равновесных сообществ. Это прежде всего экосистемы с постоянными условиями, с круглогодичным наличием пищи. Они почти не изменяются на протяжении миллионов лет, если не считать случаев замещения одних видов другими. Такой тип эволюции, протекающей в стабильных экосистемах и сильно заторможенной системой, Красилов назвал когерентной эволюцией (от лат. «cohaerens» – находящийся в связи, в согласии) (Красилов, 1969). Для ее описания и создавались классический дарвинизм, СТЭ и другие традиционные теории.
Но существует другой тип эволюции – некогерентной (там же). Эволюция такого рода протекает в неустойчивых, нарушенных экосистемах, или, что то же самое, экосистемах, находящихся в состоянии кризиса. Современную ЭТЭ, в противоположность СТЭ, интересует в первую очередь некогерентная эволюция. Сторонники этой новой модели считают (и стремятся доказать), что самые важные эволюционные события – появление эукариотической клетки, многоклеточности, полового размножения, планов строения, цветка, теплокровности, плаценты, интеллекта – совершались в периоды кризисов, в фазе некогерентной эволюции. Какие же изменения испытывали в этих условиях экосистемы и есть ли в этих изменениях какая-то общая закономерность?
Данные изменения вполне аналогичны тем, что происходят на наших глазах в современных экосистемах, разрушаемых неразумной хозяйственной деятельностью человека. Длительное воздействие нового непривычного климатического режима (в основном изменявшегося в сторону похолодания) дестабилизирует экосистемы, выводя их из состояния гомеостатического равновесия. Это пагубно сказывается в первую очередь на узкоспециализированных формах-эдификаторах. В кризисных условиях таковыми, естественно, оказываются самые приспособленные и наиболее конкурентоспособные виды климаксовой стадии. Они обладают очень тонкими адаптациями ко всему комплексу средовых условий, с изменением которых эти адаптации разрушаются. Кроме того, восстановительные процессы в экосистеме, включая сукцессии, теперь оказываются прерванными. В итоге, если прежние условия не возвращаются к норме, доминантные формы вымирают.
Летопись великих кризисов всецело подтверждает это правило. Красилов приводит далеко не полный перечень господствовавших мезозойских групп организмов, которые вымерли в конце мелового периода. Это, в частности, беннетики и чекановские среди растений, аммониты, иноцерамусы и рудисты – среди моллюсков, динозавры, птерозавры и часть сумчатых – среди наземных позвоночных (Красилов, 1985).
С вымиранием доминантов происходит упрощение структуры экосистем. В них развиваются энтропийные процессы, обратные тем прогрессивным тенденциям, которые характерны дня спокойных бескризисных периодов. Сокращаются видовое разнообразие и число пищевых цепей, прекращается сукцессия, падают продуктивность и биомасса и растет мертвая масса (Krassilov, 1994; Красилов, 2001).
Но вымирание доминантов имеет по меньшей мере одно важное эволюционное следствие – с их исчезновением освобождаются экологические ниши. Природа, как известно, не терпит пустоты, и в освободившиеся ниши устремляются виды, доселе занимавшие скромное место в экономике экосистемы. Особо отметим, что смена «владельцев» ниш происходит не насильственным путем, не в силу конкурентной борьбы и вытеснения одного вида другим, более приспособленным, как это следовало из теории естественного отбора. Все происходит по более простой и понятной схеме.
Виды, занявшие вакантные ниши, обладают типично пионерскими свойствами. Они неприхотливы, сравнительно слабо специализированны, у них короткий жизненный цикл, высокая смертность (они – объект массовой неизбирательной элиминации), но она компенсируется высокой плодовитостью. В условиях, когда в силу упрощения структуры экосистемы конкуренция (межвидовая) резко упала и интенсивность стабилизирующего отбора снизилась, виды-пионеры в состоянии выжить именно благодаря своей высокой репродуктивной потенции. Теперь после случайных локальных катастроф эти виды благодаря своим свойствам способны взять на себя функцию быстрых регенераторов сообщества и, что еще важнее, подготовить почву для появления более конкурентоспособных видов следующей стадии.
Подведем некоторые итоги и сделаем главный вывод.
Казалось бы, периоды кризисов и фазы некогерентной эволюции характеризуются одними негативными показателями: вместо выживания наиболее приспособленных происходит их вымирание, вместо естественного развития сукцессии – ее прекращение, вместо роста продолжительности жизни – ее укорочение и высокая смертность видов-пионеров. Растет производство энтропии. Все это признаки биологического регресса.
Однако именно в эти периоды закладывается фундамент будущего прогресса. В силу ослабления стабилизирующего отбора создаются условия для генетического поиска и резкого увеличения размаха изменчивости (в том числе за счет выявления ее скрытого резерва). Когда конкуренция падает, живая природа может позволить себе эволюционное экспериментирование, и палеонтологическая летопись подтверждает, что в кризисные периоды как раз и рождаются в массе жизнеспособные монстры, появление которых в фазе когерентной эволюции гораздо менее вероятно.
Кризис – это состояние среды и живых организмов, когда невозможное становится возможным. Из генетики хорошо известна сложно организованная многоуровневая система противоинформационной защиты, существующая у всех эукариотных организмов. Своей изощренностью и степенью надежности она способна поразить воображение любого человека. Оказывается, однако, что при стрессе эта система резко снижает свою эффективность и как бы сама «попустительствует» проникновению в организм экзогенного генетического материала. Явление, аномальное и вредное для вида в привычных условиях, оборачивается в частое и благое в кризисной ситуации: благодаря активизации МГЭ чужеродные гены принимают участие в желанной реорганизации его генома. И в этом аспекте, как мы видим, ЭТЭ, разработанная Красиловым, совпадает с представлениями, уже известными нам из новой генетики.
Если геологический кризис развивается медленно, то биологическая эволюция экосистем совершается быстро, иначе быть просто не может, поскольку организмы не могут длительное время пребывать в состоянии стресса: они должны или погибнуть, или найти способ измениться. В этом поиске будут задействованы не отдельные виды, а сразу большинство видов сообщества, и в результате по выходе из кризиса его состав почти полностью обновится. Фактически это будет новая экосистема.
Красилов считает ЭТЭ противостоящей неокатастрофизму, включая астероидную гипотезу. Дело в том, что адепты неокатастрофизма принимают, например, падение на Землю гигантского небесного тела и вызываемое им сильное замутнение атмосферы за непосредственную причину уничтожения видов, тогда как в ЭТЭ геологические события – всего лишь пусковой механизм в дестабилизации экосистем. Кроме того, число данных, подтверждающих ЭТЭ, с каждым годом увеличивается. С астероидной гипотезой иная ситуация. В силу недостаточной разрешающей способности существующих стратиграфических методов подтвердить ее вообще невозможно, зато опровергнуть нетрудно. В частности, все больше свидетельств того, что обнаруженные иридиевые слои в отложениях соответствующих эпох – главный козырь гипотезы – имеют скорее магматическое, чем космическое, происхождение (см. об этом в гл. 9).
Правда, надо признать, что в плане обоснованности у ЭТЭ остаются и, возможно, навсегда останутся свои узкие места. Главное из них связано с недостаточностью доказательств хронологического соответствия между действием дестабилизирующих абиотических факторов (климата, магнитного поля и др.) в периоды геологических диастроф и биотическими революциями. О трудностях точной хронологической привязки вымираний тоже уже говорилось (см. гл. 9). В этом, в частности, состоит одна из причин, почему ЭТЭ рассматривают как гипотезу.
