Глава 17
ГЕНЕТИКА И ЭПИГЕНЕТИКА ВИДООБРАЗОВАНИЯ И МАКРОЭВОЛЮЦИИ
Ушедший XX век подвел науку об эволюции к парадоксальной ситуации. Генетика с ее специализированными материальными носителями наследственности, к открытию которых упорно стремились многие поколения исследователей, те самые молекулы ДНК, которые еще недавно царствовали в умах биологов-эволюционистов как азбука любых перемен в живом, вдруг оказались малозначимыми для теории эволюции! У эволюции свои законы, и генетическая система у них всего лишь в подчинении. Зато эволюция, если говорить образно, шагает под руку с эпигенетикой и даже позволяет ей время от времени себя вести.
Между тем эпигенетика пользуется языком генетики и считает для себя необходимым изучать все, что происходит в геноме. Причина столь разного отношения эволюции и эпигенетики к генетике ясна: эволюция вполне «сознает», что генетический аппарат – ее порождение, но никак не причина. Тогда невольно возникает вопрос: зачем ученым-эволюционистам интересоваться генетическими изменениями, если они не первичны?
Вероятно, данный парадокс способен вызвать то же замешательство, какое может испытывать здравомыслящий гражданин, впервые увидевший уличный щит с рекламой сигарет, а под ней неброское предупреждение Минздрава о том, что курение опасно для здоровья,
В данном случае, как нам представляется, разумно опираться на следующее соображение. Претерпевая те или иные изменения и осуществляя генетический поиск, генетический аппарат создает базу для надежного воспроизведения тех удачных поведенческих и физиологических реакций, которые осуществил организм в ответ на сдвиги во внутренней или внешней среде.
Перед нашим взором прошло достаточно примеров крушения разных догм и «возвращения к истокам». Не будем удивляться и упомянутому парадоксу и воспримем его с надеждой, что в будущем наука сможет его удовлетворительно разрешить. Смиримся также с фактом, что при всех успехах генетики и селекции мы не можем похвастаться созданием сколько-нибудь значительного числа полноценных биологических видов. С точки зрения наших познавательных возможностей это вызывает разочарование, но зато служит дополнительным стимулом к созданию истинной и, значит, действенной теории видообразования.
Обратимся теперь к тому, что известно из области эпигенетических и генетических событий, сопутствующих образованию видов и более высоких таксонов. Только будем помнить, что генетические изменения не диктуют организму, что ему делать, а всего лишь фиксируют в своей генетической памяти его правильные эволюционные шаги.
За последние 30 лет здесь произошла еще более радикальная революция, чем в молекулярной генетике. Прежние представления о способах видообразования, основанные на модели СТЭ, оказались совершенно несовместимыми ни с теорией прерывистого равновесия, ни с эпигенетикой, ни с новой генетикой. На смену им пришло множество альтернативных гипотез, часть из которых успели обрести статус достаточно прочно обоснованных теорий.
Все эти годы объем непредвзятых исследований по проблеме видообразования продолжал нарастать, но все попытки выяснить, какие именно генетические изменения (и в каком количестве) связаны с видообразованием, до сих пор не привели к какой-либо единой схеме. И это вполне объяснимо, ибо в связи с различиями в строении генома, в детерминации и течении процессов индивидуального развития, в способах размножения и т. п. у разных групп организмов и не может быть единого механизма видообразования. Не приходится удивляться, таким образом, что органический мир демонстрирует нам множественность путей и способов образования новых видов, с некоторыми из которых мы уже знакомы.
Разнообразие, однако, не исключает принципиальной общности. Главные способы видообразования, которые мы рассмотрим ниже, отвечают следующим характеристикам.
Новые виды образуются только сальтационно в течение одного-двух или ограниченного числа поколений в результате установления репродуктивной изоляции. При такой скорости видообразовательного акта захват популяции новым видовым комплексом признаков путем вытеснения старого на основе генетико-популяционных процессов невозможен, так как такие процессы протекают слишком медленно и обычно не имеют завершения. Поскольку популяционные процессы к видообразованию не ведут, естественный отбор оказывается без объекта своего действия. К тому же возможные эффекты отбора резко подавлялись бы системой активных реакций организма. Вид рождается сразу в большом числе особей, реже – в нескольких. Изменения, осуществляемые организмом, часто носят адаптивный и направленный характер и оказываются в разных таксонах параллельными, что свидетельствует о закономерности их возникновения и справедливости номогенетической концепции. Если изменения носят иной характер, то равновесие со средой устанавливается преадаптационным путем.
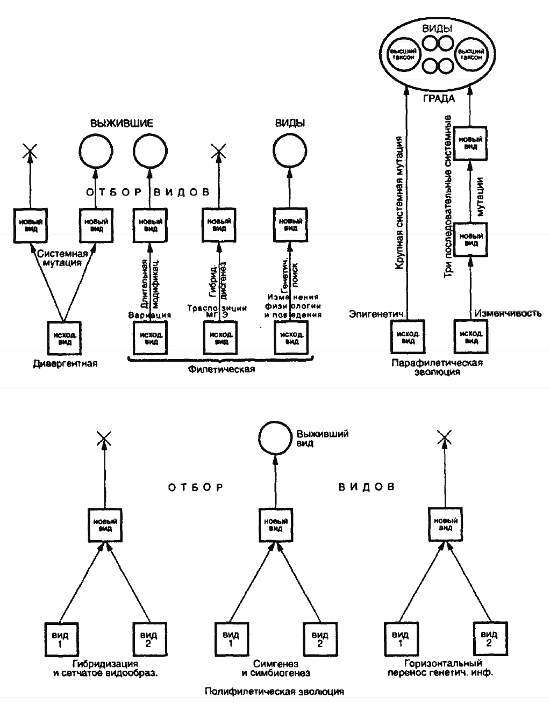
Рис. 27. Обобщенная схема, иллюстрирующая множественность путей видообразования (ориг.)
Особо подчеркнем, что приведенная характеристика видообразования, каковой она видится в свете новой генетики, всецело соответствует теории прерывистого равновесия (см. гл. 15).
В синтетической теории считается, что видообразование – сложный и длительный процесс, не сводимый к изменчивости. Последняя служит всего лишь «сырым материалом» для отбора, который выступает единоличным творцом новых форм. Все попытки при объяснении видообразования ограничиться одной изменчивостью, проигнорировав отбор, адептами СТЭ всегда резко осуждались. Сейчас нам совершенно ясно, почему механизмы формообразования нельзя, с позиций СТЭ, свести к изменчивости: механизм естественного отбора и целесообразные действия организма – это разные и несовместимые способы эволюции. При первом способе изменчивость безлика (неопределенна), при втором – выражение целесообразных реакций.
В новых теориях и гипотезах видообразования, как правило, обходятся без отбора и действительно сводят этот процесс к наследственной изменчивости. При этом авторы теории вовсе не считают изменчивость простым процессом. Мы убедились, что ей могут соответствовать сложные генетические и эпигенетические процессы и, как считают некоторые специалисты, и она может быть результатом своеобразных внутригеномных популяционных взаимодействий. Короче, нет ничего научно порочного или методологически предосудительного, чтобы считать носителей вариационной или динамической изменчивости новыми видами, если они этого заслуживают.
