Взаимодействие ОК – ФК и две формы наследственной изменчивости
Облигатный и факультативный компоненты генома не изолированы друг от друга. И самое интересное – это взаимодействие ОК и ФК, постоянная и относительно свободная миграция генетических элементов между ними. Как они осуществляются?
Переход ОК в ФК может происходить в процессе амплификации генов, в том числе путем их захвата ретровирусами. Примером обратного перехода ФК в ОК могут служить так называемые инсерционные мутации – продукт внедрения самых разных элементов ФК, способных к взаимопревращениям. Голубовский особенно акцентирует внимание на вирусах (ретровирусах), которые, как многие полагают, способны выступать не только в роли усилителя мутационного процесса, но также в качестве доноров и переносчиков мобильных генетических элементов, осуществляющих генетический обмен между разными видами (о горизонтальном переносе генов мы уже говорили в гл. 12).
Одним из наиболее изученных в генетическом отношении и показательных примеров взаимодействия двух наследственных систем могут служить сложные морфофизиологические наследственные изменения генотипа дрозофилы, вызванные присутствием вируса сигма. Еще в 1937 г. Ф. Л'Эритье обнаружил у дрозофилы мутацию, вызывавшую гибель мутантных мух в атмосфере углекислого газа. Тогда было с удивлением отмечено, что мутация наследуется не по закону Менделя, а через цитоплазму, причем как по материнской линии, так и через самцов. Это явление долгое время оставалось загадкой. Только спустя много лет удалось выяснить, что причиной необычного наследования оказался РНК-содержащий вирус, размножавшийся в половых и соматических клетках мутантных мух и служивший источником их чувствительности к углекислому газу (L'Heritier, 1970; см.: Brun, Plus, 1980; Landman, 1991). У этого вируса были также найдены мутации, при которых скорость его репликации заметно возрастала, и тогда он (вместе с признаком чувствительности к СО2) начинал передаваться и через самцов. Если температуру содержания мух повышали до 30 °С, происходило их освобождение от вирусов и, соответственно, исчезал также признак чувствительности к СО2. Это приобретенное мухами в ходе онтогенеза свойство сохранялось (наследовалось) в ряду многих поколений. Было также установлено, что в природных популяциях носителями вируса оказывается до 20-50% особей.
Разделение генетической системы эукариот на ОК и ФК привело Голубовского к новым, расширенным представлениям о наследственной изменчивости. Уже в 1978 г. он высказал принципиально важное предположение, что взаимодействие ОК и ФК служит основным источником наследственных изменений в природе. В дальнейшем (Голубовский, 1985а, б) он выделил три формы наследственной изменчивости: мутационную, вариационную и динамическую. Первые две связаны с изменением структурных компонентов генома, третья – с особыми регуляторными механизмами его оперативной памяти. Все три формы изменчивости изучались автором на природных популяциях Drosophila melanogaster в течение 20 лет. Остановимся сначала на двух первых.
Изменения, прямо или косвенно затрагивающие ОК генотипа – структуру генов, их расположение в хромосоме и число хромосом,– это обычные канонические мутации, принятые в классической генетике. К ним относятся, в частности, наследственные изменения, получаемые в лаборатории, которые связывают с «поломками» ДНК. Мутации – это лишь часть существующей в природе изменчивости и далеко не самая важная.
Со структурными изменениями ФК связана специфическая форма изменчивости, которую Голубовский предложил именовать вариационной. Особенности мутаций хорошо известны: они возникают случайно, спорадически и с относительно низкой частотой. Характер вариаций совсем иной. При них часто наблюдаются массовые, упорядоченные наследственные изменения, на которые в свое время указывал Берг, а затем его последователи и единомышленники. Вариации обычно возникают при таких изменениях абиотической, биотической или генетической среды, при которых канонические мутации наблюдаются редко.
На схеме, приводимой Голубовским, представлены взаимодействия среды и двух компонентов генотипа в виде следующей триады (рис. 26). На ней толщина стрелок отражает степень влияния одного члена триады на другой.
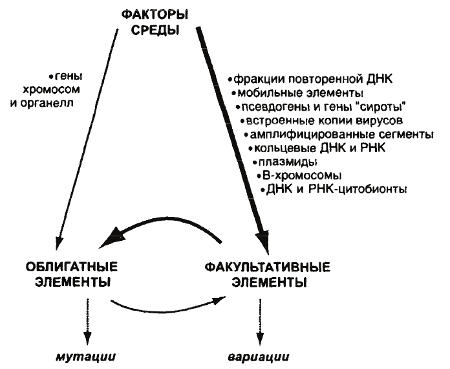
Рис. 26. Пути возникновения наследственных изменений в природе в системе «среда-факультативные-облигатные элементы» (из: Голубовский, 2000)
В классической генетике связь среды с ОК принималась, как правило, за единственную или доминирующую форму наследственной изменчивости. Обнаружилось, однако, что в природе существуют более мощные потоки, приводящие к генетическим изменениям: «среда→ФК» и «ФК→ОК». Именно эти потоки служат основным источником наследственных изменений вида в природной обстановке. В свете данного открытия различия между изменениями, получаемыми у лабораторных животных с помощью искусственных мутагенов, и изменениями, наблюдаемыми в естественных популяциях, получают прочное обоснование, и это весьма существенно как для генетики, так и для эволюционной теории. В этой связи вспомним новосибирского генетика Ю.Я. Керкиса (1940). В далеком 1940 г. он в числе немногих осознал это различие и высказал правильную догадку, что спонтанные наследственные изменения возникают вследствие нарушения внутриклеточного метаболизма и физиологического гомеостаза. Именно эти причины в первую очередь вызывают изменения факультативных элементов, а затем – опосредованно – и ОК.
В вариационной изменчивости особенно важны количественный состав ФК и процентное отношение его фракций к ОК, которое у разных видов нередко поддерживается на постоянном уровне. В случае генетического стресса, когда активность генов в ФК повышается, происходит магнификация генов и возрастает вероятность включения их дополнительных копий в ОК со всеми вытекающими отсюда эволюционными следствиями.
