Но вернемся к существу пунктуализма. Эта концепция сделала видообразование центральным событием в структуре эволюции и основным звеном в макроэволюции. Видообразование имеет всегда квантовый характер и происходит скачком: «…мы должны отвергнуть градуализм как ограничительную догму. Пунктуационное изменение с резкими скачками между стабильными состояниями характерно для большей части нашего мира»,– заявляет Гулд (1986. С. 39). Его мнение разделяют большинство крупнейших генетиков: «…эпизоды видообразования включают в себя … значительные генетические скачки, так что формирование новых видов осуществляется в серии катастрофических, стохастических генетических событий» (Carson, 1975. Р. 87-88).
Продолжительность скачка может быть самой разной – от превращения вида в одном поколении до ступенчатых преобразований, растягивающихся на несколько десятков тысяч лет. Вероятно, для пунктуализма в большей мере характерно признание первичности, исходности репродуктивной изоляции, которая рассматривается не как продукт адаптации, а как стохастическое явление. В этой связи Гулд (Gould, 1980), например, считает, что традиционные понятия алло- и симпатрического видообразования утратили свое значение. Если демы оказываются разобщенными, новые виды могут возникать в любой точке ареала предкового вида. Вместе с тем очевидно, что само видообразование становится неадаптивным стохастическим процессом, практически совершающимся независимо от естественного отбора.
Масштабы видообразовательных актов и способы их реализации, допускаемые «прерывистой» концепцией, достаточно разнообразны. Внезапно возникающие новые формы могут не выходить за рамки видового статуса, и тогда, согласно Гулду, анагенез оказывается просто «аккумулированным кладогенезом» (Gould, 1982a. Р. 139). Но Стэнли (Stanley, 1979) допускает и даже считает правилом, возникновение родов и более высоких таксонов «главным образом благодаря квантовому видообразованию», и притом в течение нескольких поколений. Так, он пишет: «Чтобы возникли новые роды, подсемейства и семейства, достаточно единственного акта видообразования или краткой последовательности событий» (ibid., p. 141). Однако главный механизм макроэволюции, по Стэнли, иной, и о нем будет сказано отдельно.
Карсон (Carson, 1975, 1978) разработал оригинальную теорию («founder-flush-theory») чрезвычайно быстрого видообразования, опираясь на принцип основателя и генетический дрейф. По этой теории видовые популяции последовательно переживают состояния взрыва («flush»), когда внезапно возникает много особей, и крушения («crash»), когда большинство их гибнет. Единичные выжившие демы в предельном случае могут состоять из одного индивида-основателя, претерпевшего вынужденную генетическую реорганизацию. Это так называемый диахронный эффект бутылочного горлышка. По Карсону, он составляет сущность процесса видообразования (Carson, 1975. Р. 88) (рис. 25). Стэнли трактует этот способ как «потенциальный механизм ускорения филетической эволюции», родственный действию катастрофического отбора, и на этом основании (ввиду «чрезвычайной редкости») фактически его отвергает (Stanley, 1979; Р. 98). Действительно, при данном способе увеличения числа видов не происходит, а факты длительного существования хроновидов заставляют признать его весьма ограниченную роль в макроэволюции. Стэнли отстаивает ключевую роль в макроэволюции очень малых популяций, оказавшихся просто изолированными от нормальных популяций предшествующих видов, что и следует считать типичным для гипотезы прерывистого равновесия. Вместе с тем он положительно относится к идее возникновения нового вида (например, у брюхоногих моллюсков) от единственной гермафродитной самки, испытавшей зародышевую мутацию.
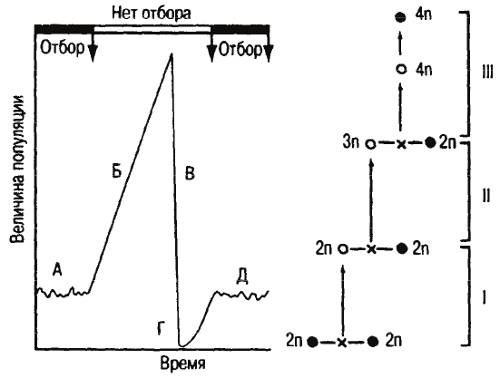
Рис. 25. Схема видообразования по Карсону (1975)
Пунктуализм полностью восстановил в правах идею Гольдшмидта о макроэволюционной роли «обнадеживающих уродов» и фактически дал ей новую жизнь (Steenis, 1969; Frazzetta, 1970; Bush, 1975; Gould, 1977b; Stanley, 1979). Теоретики пунктуализма Гулд и Стэнли лишь формально делают оговорки о «крайности» и «фантастичности» этой идеи, но принимают ее по существу. Констатируя возвращение гипотезы Гольдшмидта об «уродах», Буш резюмировал, что она «больше не является полностью неприемлемой» (Bush, 1975. Р. 357). Главное затруднение, с которым гипотеза встречалась еще в 60-х – начале 70-х годов,– проблематичность нахождения уродом пары для производства плодовитого потомства – ныне полностью преодолено. Уже тогда было установлено, например, что резко аберрантная форма способна закрепиться, если она происходит через зародышевую мутацию самки, как это было показано в упоминавшихся работах Карсона о видообразовании у дрозофил на Гавайских островах.
Пунктуализм принял и другое положение Гольдшмидта – о полном разобщении микро- и макроэволюции, которое теперь именуется разрывом Гольдшмидта (Gould, 1982a. Р. 137), или главным положением теории прерывистого равновесия (Maynard Smith, 1982. P. 126). Считается, что Гольдшмидт предугадал прерывистое видообразование и его кладистический характер.
Действительно, принцип разобщенности микро- и макроэволюции в начальном варианте «прерывистой» концепции – основной, определяющий ее структуру и само содержание. Обосновывается он, как и в свое время Гольдшмидтом, приведением в действие в случаях микро- и макроэволюции различных типов изменчивости и механизмов ее осуществления. Макроэволюционное значение внутривидовой изменчивости и полиморфизма, как правило, отрицается. К природе изменчивости и генетике видообразования вообще проявляется особый интерес. Но одним фактором изменчивости дело не ограничивается. При объяснении макроэволюции не менее важен отбор, только он перенесен на другие уровни организации.
Один из веских доводов за разобщенность микро- и макроэволюции – зависимость их темпов от разных причин, подмеченная многими специалистами. Вспомним основополагающую работу Симпсона (1948). В ней ясно показано, что ни одна из причин, обычно определяющих темп микроэволюции, на скоростях макроэволюции не сказывается. Можно привести показательный пример. Род дрозофилы, появившийся в эоцене, хотя и представленный многочисленными видами (с вымершими – более двух тысяч), несмотря на быструю смену генераций и высокую насыщенность популяций мутациями, так и не дал нового рода, тогда как у хоботных, размножающихся на несколько порядков медленнее и едва ли столь насыщенных мутациями, за то же геологическое время сменилось несколько родов. Следовательно, макроэволюция у хоботных проходила в несколько раз быстрее, чем у дрозофил.
Как свидетельствуют данные палеонтологии, темпы макроэволюции зависят от таких показателей, как размер популяции, частота видообразования, изменение общей биоценотической обстановки. Отсюда закономерен вывод, что различие причин, от которых зависят темпы микро- и макроэволюции, говорит об отличии на этих уровнях движущих сил эволюции.
