Симбиогенез
Кроме гибридогенеза в природе существует и такой способ объединения разнородных организмов с образованием новых, при котором слияния геномов исходных форм не происходит. В результате образуются организмы-кентавры, классическим примером которых могут служить лишайники.
Впервые мысль о двойственной природе этих растений была высказана швейцарским ботаником С. Швенденером (Schwendener, 1869). В начале 1880-х годов она получила полное подтверждение. Было доказано, что лишайники представляют собой продукты эволюционного объединения гриба и водоросли, т.е. получилось, что целый отдел царства растений возник не путем дивергенции, а с помощью обратного процесса – слияния ранее совершенно самостоятельных организмов.
Однако, как показала Л.Н. Хахтна (1973, 1975, 1979), заслуга в изучении этого удивительного феномена, поразившего многих современников, в гораздо большей степени принадлежит русским ботаникам, которые в ряде моментов даже чуть опередили своего швейцарского коллегу.
В 1867 г. А.С. Фаминцын и О.В. Баранецкий опубликовали на немецком языке (Famintsin, Baranetsky) сообщение об опытах, в которых им удалось отделить гонидии (одноклеточные зеленые водоросли) лишайника от его бесцветного слоевиша. Гонидии оказались способными к самостоятельному существованию в куль-туре, подобно своим свободноживущим собратьям, образовывали споры и по строению весьма походили на таковых рода Cystococcus. Из соображений научной щепетильности авторы еще не заявили о двухкомпонентной природе лишайника, они воздержались от принятия этой идеи и тогда, когда ее высказал Швенденер, но сделанное ими открытие фактически уже означало положительное решение этого вопроса.

Андрей Сергеевич Фаминцын (1835-1918)
Через 40 лет А.С. Фаминцын обратился к проблеме роли симбиоза в эволюции и в серии статей (1907а, 1907б, 1912а, 1912б и позднее) попытался представить симбиоз как важный формообразующий фактор, дополняющий работу дивергентного видообразования. Интересно, что теперь Фаминцын (1907) задался идеей показать, что принцип, породивший лишайники, можно распространить и на растительную клетку как структурную единицу.
Чуть раньше с аналогичной гипотезой – независимо от Фаминцы на – выступил ботаник Казанского университета К.С. Мережковский (Mereschkovsky, 1905), брат поэта Д.С. Мережковского. Так родилось учение о симбиотическом происхождении клетки зеленых растений, которое после введения Мережковским в 1909 г. ныне общепринятого термина «симбиогенезис» (симбиогенез) стало называться учением о симбиогенезе. В переводе с греческого это слово означает «возникновение на основе совместной жизни».
По мнению обоих авторов этой фантастической гипотезы, и ядро, и хлоропласты (хроматофоры) с заключенным в них хлорофиллом, и центросомы – короче, все известные тогда органеллы растительной клетки – ведут свое происхождение от бактерий и водорослей, которые проникли некогда в бесцветный амебоподобный (или флагеллятоподобный) животный организм извне и стали его постоянными симбионтами. При этом особое значение для подтверждения гипотезы имели хлоропласты с их большой автономией и, как были убеждены авторы, непрерывностью этих пластид в ряду клеточных поколений; важным представлялось их сходство с современными свободноживущими одноклеточными водорослями.
Фаминцын полагал, что хлоропласты ведут свое начало от таких форм, как хлореллы и ксантеллы. Мережковский вел их генеалогию от примитивных сине-зеленых (цианей). При этом оба категорически отвергали традиционные представления об их образовании всякий раз заново путем дифференциации клеточной плазмы.
Надо сказать, что Мережковский шел в своих рассуждениях дальше Фаминцына. Он был убежден (Мережковский, 1909), что в основе всего живого лежат две глубоко различные плазмы – микоидная, из которой состоят бактерии, сине-зеленые водоросли и большая часть грибов, и амебоидная, которая слагает ткани животных и растений. Организмы, составленные из микоидной плазмы (Мережковский называл их микоидами), принадлежат к самому древнему царству на Земле. В результате симбиоза простейших безъядерных амебоидных существ (монер) с первичными микоидами – бактериями биококками – возникли первичные одноклеточные организмы – амебы и флагелляты. Бактерии образовали ядро клетки. Затем благодаря симбиогенезу совершился новый «творческий акт»: в первичные амебы и флагелляты внедрились сине-зеленые водоросли и превратились в хлоропласты. Так, путем двойного симбиоза возникли клетки всех высших растений.
К концу 1930-х гг. интерес к проблеме симбиогенеза, столь активно обсуждавшейся до сих пор как в отечественной, так и в зарубежной науке, резко падает, а вскоре о ней и вовсе забывают. Это объясняется в основном двумя причинами. С исчерпанием возможностей светового микроскопа в исследовании микроструктур клетки дальнейшие дискуссии становились бесплодными, а все попытки (в том числе Фаминцына) культивирования в искусственных средах зерен хлорофилла, извлеченных из растительной клетки, оказались тщетными.
Но из теоретических построений Мережковского по крайней мере одна идея – о двух плазмах – оказалась пророческой. Начиная с 60-х годов в сознании цитологов и микробиологов все более крепло представление о глубокой пропасти, которая разделяет безъядерные (точнее было бы сказать – доядерные) организмы, каковыми являются бактерии, и ядросодержащие, к которым относятся все остальные. Коренные различия между ними, как выяснилось, распространялись и на ультраструктуры, в том числе формы укладки ДНК (Кернс, 1967). В итоге оказалось, что различия между ядерными и безъядерными глубже и фундаментальнее, чем между традиционными царствами животных и растений, и что вполне обоснованно делить все живое на два надцарства – прокариот и эукариот (Тахтаджян, 1973). Замечательно, что границы этих надцарств (за исключением грибов) совпали с распределением организмов между двумя типами плазмы у Мережковского.
Успехи в изучении ультраструктур клетки 1960-х гг., ставшие возможными благодаря новым методам исследования (электронная микроскопия, центрифугирование, усовершенствование биохимических, цитофизиологических и других методов), создали основу для возрождения гипотезы Фаминцына – Мережковского на новом уровне. С обновленной гипотезой о симбиогенетическом происхождении эукариотической клетки выступила молодой биолог из Бостонского университета Линн Саган-Маргулис (Sagan, 1967; Margulis, 1970 и позднее). Не упомянув своих русских предшественников, она развила ряд положений, очень близких представлениям Мережковского.

Линн Маргулис (род. 1938)
Л. Маргулис опиралась теперь на целый ряд гораздо более достоверных фактов, свидетельствовавших об автономии клеточных органелл, их сходстве друг с другом, а также с цианеями и бактериям. Было установлено, что как пластиды, так и митохондрии способны к авторепродукции, причем не всегда синхронизированной с митотическим циклом клетки. Они содержат собственный генетический аппарат, в значительной мере автономный от ядерной ДНК, причем на долю внеядерной ДНК приходится от 5 до 40% всей ДНК клетки. Обнаружилось также, что ДНК органелл всех до сих пор изученных эукариотических организмов сложена в виде колец, как это наблюдается у всех бактерий и одноклеточных сине-зеленых водорослей, тогда как ядерная ДНК эукариот образует хромосомы, располагающиеся линейно (Кернс,
1967). Сообразно генетическим различиям пластиды и митохондрии обладают и своим высоко-автономным белоксинтезирующим аппаратом (Филиппович, Светайло, Алиев, 1970).
Зато по тем же и целому ряду других характеристик хлоропласты оказались сходными с сине-зелеными водорослями (Taylor, 1970; Пахомова, 1972, 1974), а митохондрии – с бактериями (Nass, 1969; Рудин и Уилки, 1970).
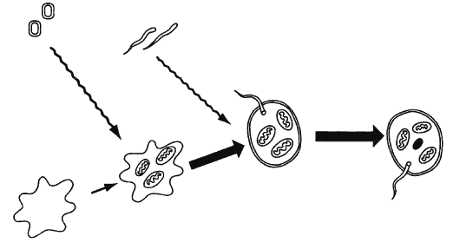
Рис. 13. Схема симбиотического возникновения эукариотической клетки по Маргулис (из: Тахтаджян, 1973)
Опираясь на эти данные, Маргулис (Margulis, 1970) предложила следующую модель симбиотического возникновения эукариотической клетки (рис. 13). Родоначальником всех форм жизни, от которого не менее чем 3,3 млрд лет назад началась органическая эволюция на Земле, был небольшой гетеротрофный амебоидный прокариотный организм, еще не способный дышать кислородом. Эти гипотетические допотопные организмы поглотили, не убивая, более мелкие аэробные бактерии, которые в теле своих хозяев превратились в митохондрии (первый этап симбиогенеза). Образовавшиеся в результате более крупные микоплазмоподобные организмы приобрели высокоподвижные спирохетоподобные бактерии и стали жгутиконосными формами. Спирохетоподобные бактерии способствовали образованию настоящего ядра, жгутикового и митотического аппаратов – возникли простейшие эукариотические организмы, давшие начало царствам животных и грибов (второй этап симбиогенеза). Наконец, последним этапом эволюции эукариотической клетки стало объединение с фотосинтетиками типа примитивных цианей, которые превратились в фотосинтезирующие пластиды и открыли своим хозяевам путь к приобретению автотрофного типа питания (третий этап симбиогенеза). Этот последний этап дал начало всему стволу растений. Таким образом, все эукариоты, по Маргулис, являются по меньшей мере двухгеномными организмами.
Симбиогенетическая гипотеза возникновения эукариот получила широкую поддержку и распространение как на Западе (Raven Р.Н.; Schnepf E., Brown R.M.; см. сводку: Cavalier-Smith, 1995), так и в России (Тахтаджян А.Л.; Генкель П.А.; Яблоков А.В. и Юсуфов А.Г.; см. обзор: Кусакин, Дроздов, 1994). В своей последней книге по эволюции Н.Н. Воронцов заявил, что он принимает «гипотезу симбиогенеза как наиболее вероятную» (Воронцов, 1999. С. 496). При этом он справедливо добавляет, что последовательность этапов симбиогенеза могла быть и другой, но в любом случае эволюционное развитие не могло быть монофилетическим. Впрочем, есть исследователи, не согласные с Маргулис. Они считают, что митохондрии и хлоропласты не произошли от бактерий, а всего лишь получили от них ряд генов (Gogarten, 1995).
Таким образом, как и в случае гибридогенеза и полиплоидии, когда образование новых форм происходит благодаря слиянию и дупликации геномов, возникновение новых таксонов путем симбиогенеза связано с пространственной интеграцией двух или более геномов в рамках одного организма (его клеток), т.е. осуществляется путем синтезогенеза, прямо противоположным дивергенции. Здесь нет места для каких бы то ни было процессов микроэволюции. Это чисто макроэволюционный (точнее, мегаэволюционный) процесс.
Обсуждение проблемы симбиогенеза и всех новейших открытий молекулярной биологии, на которые она опирается, побудило многих исследователей к перестройке общей филогенетической системы органического мира, а именно ее самых верхних этажей, составляющих предмет мегасистематики. Характерной тенденцией последнего времени стало увеличение числа ее высших подразделений – надцарств, царств, подцарств и типов (отделов). Так, в генеральной системе А.Л. Тахтаджяна (1973), пользующейся широким признанием, два надцарства, четыре царства и девять подцарств. О.Г. Кусакин и А.Л. Дроздов (1994) предложили систему, состоящую из 22 царств и 132 типов. Интерес к соотношению объемов высших таксонов сохраняется по сей день.
