Одна из первых догадок о существовании полиплоидии и гибридизации у позвоночных принадлежит шведскому зоологу Г. Свердсону (Svardson, 1945). По его мнению, таким путем могло возникнуть семейство лососевых рыб. Кариологические исследования и данные электрофореза, полученные в 1970-е годы B.C. Кирпичниковым (1979) и Ю.П. Алтуховым (1989), подтвердили это предположение и выявили другие группы рыб, в эволюции которых гибридизационные процессы играли важную роль. Полная сводка всех известных фактов этого рода содержится в книге В.П. Васильева (1985).
Л.Я. Боркин и И.С. Даревский (1980) описали неординарный тип гибридогенного видообразования у ряда амфибий (роды Ambystoma и Rana) и рептилий (роды Lacerta и Chemidophorus). Они, в частности, указали, ссылаясь на работы Л. Бергера (Berger) и других исследователей, что обычная прудовая лягушка Rana esculenta представляет собой гибридную форму от скрещивания R. lessonae и R. ridibunda.
Эти авторы изложили концепцию последовательной гибридизации с обязательной промежуточной фазой образования диплоидной гибридной клональной и, как правило, однополой формы. Эту концепцию ныне разделяют большинство специалистов. Она предполагает существование как минимум трех основных генетических этапов гибридогенного видообразования, ведущих к аллотетраплоидии у позвоночных.
На первом этапе в результате гибридизации на уровне бисексуальных нарождающихся диплоидных видов может образоваться диплоидная форма, переходящая к клональному размножению путем гиногенеза (рыбы), «одалживания» отцовского генома на одно поколение (рыбы, бесхвостые амфибии) или партеногенеза (хвостатые амфибии, рептилии). На втором этапе вследствие возвратной гибридизации этой диплоидной однополой формы с одним из родительских видов может возникнуть триплоидная однополая форма, размножающая с помощью гино- или партеногенеза. Наконец, на третьем этапе в результате скрещивания этой последней с одним из близких бисексуальных диплоидных видов может появиться тетраплоидная форма, способная вернуться к нормальному бисексуальному размножению (рис. 11).
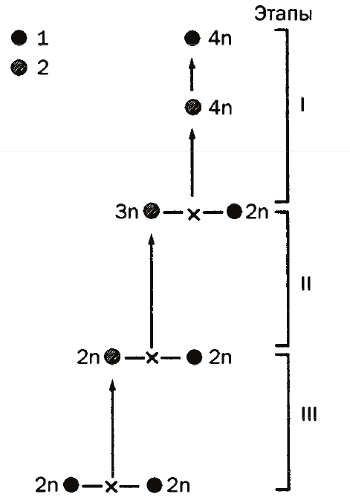
Рис. 11. Схема гибридизации (Боркин, Даревский, 1980)
Авторы отмечают, что эта схема (второй и третий этапы) хорошо согласуется с экспериментально подтвержденной гипотезой Б.Л. Астаурова (1969) о непрямом развитии естественной полиплоидии у шелковичного червя.
Для биолога должен составить особый интерес феномен «одалживания» генома самцов: он сливается с геномом гибридных самок в процессе оплодотворения, но элиминируется у их потомков в ходе мейоза. В последнее время этот феномен стали обнаруживать также в развитии некоторых гибридных беспозвоночных.
В ходе дальнейшего изучения мировой фауны ящериц общее число видов гибридогенного происхождения из разных семейств увеличилось до 40 (Даревский, 1995). Обнаружены они в самых различных частях земного шара. Таковы 14 видов североамериканского рода Cnemidophorus, комплекс видов западноавстралийских гекконов Heteronotia binoei, гекконы сборного вида Lepidodactylus lugubris с тихоокеанских островов, 5 кавказских партеногенетических видов скальных ящериц рода Lacerta, а также вьетнамская триплоидная ящерица Leiolepis guntherpetersi. Все они появились сравнительно недавно – в голоцене или плейстоцене.
Достаточно подробно изучены скальные ящерицы горного Кавказа (Даревский, 1967; Даревский, Гречко, Куприянова, 2000). Из 18 обитающих здесь видовых форм 7 представлены партеногенетическими самками, предположительно образовавшимися в результате серии актов естественной межвидовой гибридизации. Самки этих видов, имеющие диплоидный набор хромосом (2п = 38), легко спариваются с самцами бисексуальных видов, включая в свой геном мужской гаплоидный набор хромосом. В результате их потомство становится триплоидным (3n = 57), и в нем сочетаются признаки материнского и отцовского видов (с преобладанием материнских). Триплоидные формы размножаются исключительно партеногенетически. В смешанных гибридных популяциях на их долю приходится иногда до 10–12% от общей численности особей. Как и все нечетно-полиплоидные организмы, они лишены эволюционной перспективы.
Автор (Даревский, 1995) полагает, что партеногенетические виды ящериц возникали в истории их рода многократно путем скрещивания одних и тех же родительских пар. Некоторые однополые виды представляют собой отдельные клоны или совокупность немногих клонов, берущих начало от одной или нескольких самок-прародительниц. Существование партеногенетических форм дает виду биологическое преимущество: благодаря тому, что потомство оставляют все особи популяции, темп размножения вида удваивается, и это способствует поддержанию его численности на высоком уровне.
Плодовитые гибриды, встречающиеся в природе среди млекопитающих, были изучены, в частности, Н.Н. Воронцовым. В последней книге (Воронцов, 1999) он привел случаи скрещивания между малым (Spermophilus pygmaeus) и крапчатым (Sp. suslicus), между большим (Sp. major) и краснощеким (Sp. erythrogenys) сусликами.
Еще три десятилетия назад Э. Майр специально отмечал, что гибридогенное видообразование хотя и часто постулировалось, но ни разу не было доказано. Сейчас можно с уверенностью заявить, что оно точно доказано всей совокупностью цитогенетических, биохимических, иммунологических и морфологических методов.
Из всего сказанного в этом разделе следует, что рассмотренные факты гибридогенного формообразования демонстрируют принципиально недарвиновский механизм видообразования: во всех описанных случаях происходит не разделение (дивергенция) филетических линий, а их схождение. Схематически этот процесс можно отобразить в виде сетки, совершенно аналогичной той, которая возникает при построении полной генеалогии любого человеческого рода. Отсюда второе название гибридогенного формообразования – сетчатое, а в английском звучании – ретикулярное видообразование (или эволюция) (рис. 12). Этот термин ввел в биологию Ф.Г. Добжанский (1937).
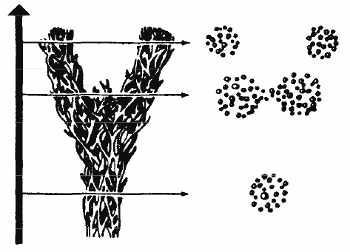
Рис. 12. Схема ретикулярной эволюции (из: Dobzhansky, 1951)
