2.5. СТРУКТУРА ПОПУЛЯЦИИ
Популяция имеет, как правило, высокодифференцированную структуру, которая выражается в существовании физиологически и морфологически различающихся групп особей с присущими им функциями. Дифференциация популяционной структуры базируется на генетической основе и направлена на максимально эффективное использование ресурсов среды. Она включена в механизм регулирования таких важных популяционных параметров, как численность и плотность (число особей на единицу площади).
Структура популяций животных. К видам, у которых популяционная дифференциация выражена слабо, относятся так называемые дисперсные виды. Помимо уже упомянутых видов птиц с характерным для них широким разлетом молодняка дисперсность присуща многим видам морских беспозвоночных. Особи этих видов на личиночных стадиях пассивно разносятся океаническими течениями на огромные расстояния, так что выделение у них местных популяций утрачивает смысл.
Вместе с тем для большинства позвоночных и для многих беспозвоночных животных формирование популяций с высокодифференцированной структурой является условием их существования.
Достаточно упомянуть общественных насекомых — муравьев, ос, пчел, термитов, у которых «разделение труда» между разными группами особей в семьях и колониях выражено исключительно ярко.
Дифференциация популяционной структуры приводит к оптимальному использованию территории и имеющихся ресурсов. Распределение особей в пространстве (пространственная структура) и система их взаимоотношений (этологическая структура) связаны. Поэтому предпочитают говорить о единой пространственно-этологической структуре, которая у каждого вида характеризуется своими особенностями. И. А. Шилов (1977) выделяет два типа таких структур. Первый тип свойствен видам оседлых животных, интенсивно использующих ресурсы территории. Второй тип присущ кочевым животным, ведущим групповой образ жизни со слабой привязанностью к территории и экстенсивным ее использованием. Между этими двумя типами имеется ряд переходных форм.
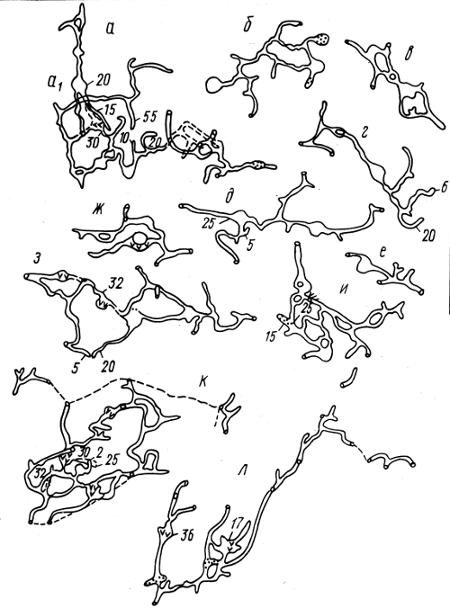
Рис. 2.14. Топография гнездовых нор на семейном участке обыкновенной полевки Microtus arvalis (по Н.В. Башениной, 1977): буквами обозначены разные типы нор, цифрами — их глубина, см
У оседлых животных, ведущих семейный образ жизни, пространственная структура реализуется путем формирования индивидуальных (семейных) участков обитания длительного использования. Распределение этих участков приурочено к определенным элементам ландшафта. Животные, овладевающие территорией, осваивают ее, что выражается в сооружении убежищ, гнезд, запасании кормов, прокладывании троп и т.д. (рис. 2.14). Обитатели такого участка безошибочно на нем ориентируются и быстро находят убежища от хищников и непогоды.
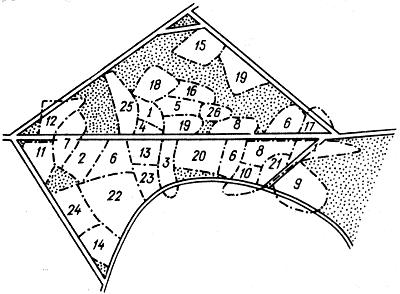
Рис. 2.15. Карта индивидуальных участков в популяции пеночки-веснички Phylloscopus trachilus (по D. May, 1949): цифрами обозначены индивидуальные участки
На рис. 2.15 дана карта индивидуальных участков в популяции пеночки-веснички. Эти участки захватываются отдельными самцами ранней весной, после чего сюда привлекаются самки и образуются семейные пары. Самцы активно защищают свои участки и, как правило, выходят победителями в случае агрессии со стороны других самцов. Наличие участка оказывается необходимым условием для нормального протекания полового цикла, гнездостроения и оставления потомства. У нетерриториальных особей половой цикл приостанавливается и они тем самым устраняются от размножения.
Из приведенных примеров следует, что индивидуальный участок представляет собой территорию, владельцы которой пользуются явным преимуществом в отношении доступа к ограниченным ресурсам среды и возможности оставления потомства.
Формы территориальности могут быть различными. Так, у бентосных рыб, древесных ящериц, многих видов птиц и млекопитающих животные на охраняемом участке добывают большую часть пищи, строят убежище, спариваются и выращивают потомство. У козодоев и тростниковых камышовок индивидуальный участок служит прежде всего для размножения, тогда как кормление осуществляется за его пределами. У многих колониальных птиц охране подлежит небольшая область вокруг гнезда.
У многих видов птиц из отряда куриных существуют участки, специально предназначенные для спаривания, — токовища. У полынного тетерева еще до сезона размножения выделяются самцы, доминирующие над другими самцами, т.е. подавляющие их активность. Они занимают на току свои индивидуальные участки. Подчиненные особи, субдоминанты, располагаются на периферии токовища. Большинство же самцов, среди которых значительная доля молодых, вообще не участвуют в токовании. Прилетающие самки в подавляющем большинстве случаев спариваются с доминантными самцами, хозяевами токовища. Последние составляют около 1% от общего числа самцов в популяции, однако на их долю приходится 80% спариваний (J. Scott, 1942; R. Wiley, 1973). Подобные же наблюдения сделаны у дымчатого, степного и других видов тетеревиных.
У шотландской куропатки, грауса, самцы занимают индивидуальные участки на вересковых болотах еще с конца лета, т.е. задолго до сезона размножения. У этого вида охраняемые участки служат как для кормления, так и для размножения и используются парой особей — самцом и самкой. В урожайные годы территориальными оказываются около половины половозрелых особей старших возрастов. Когда же случается неурожай вереска, размеры индивидуальных участков увеличиваются и соответственно резко сокращается доля территориальных особей. Птицы, оставшиеся без территории, почти все погибают в осенне-зимний период. Те же, которым удается перезимовать, имеют недоразвитые гонады и в размножении не участвуют (A. Watson, 1970).
В приведенных исследованиях, выполненных в полевых условиях, связь между пространственно-этологической структурой популяции, различиями в выживаемости особей и их успехом в размножении прослеживается очень четко. Отчетливая зависимость между зоосоциальным доминированием в стаде и частотой оставляемого потомства продемонстрирована и на домашних курах. Само это явление, называемое еще зоосоциальной иерархией, впервые было продемонстрировано на данном объекте (Th. Schjelderup-Ebbe, 1922). Такие же закономерности установлены в самых различных группах животных от беспозвоночных (некоторые крабы, раки, тараканы) до высших позвоночных, включая приматов.
У млекопитающих формируются внутрипопуляционные группировки, которые носят разные названия: демы у мышевидных грызунов, прайды у львов, гаремы у ушастых тюленей, стаи у волков и шакалов, поды у дельфинов-косаток, паки у гиеновых собак, стада у копытных, труппы у макак. В последнее время их все чаще обозначают общим термином «дем» (от греч. demos — народ). Отношения особей в таких внутрипопуляционных группировках принимают характер доминирования—подчинения и частично или полностью заменяют системы территориальных отношений, рассмотренных выше.
Домовые мыши образуют демы в замкнутых пространствах человеческих построек. В них формируется сложная структура, включающая множество групп со своей жесткой иерархией. Выделяются лидирующие самцы старших возрастов, которые резко подавляют активность всех прочих самцов в группе. Острота конкуренции непосредственно зависит от наличия ресурсов — воды, пищи, укрытий — и от плотности населения в демах. Молодые животные в основной своей массе выселяются из демов и, как правило, гибнут. Польский эколог К. Петрусевич при исследовании рыжих полевок на опытном участке леса показал, что именно мигранты, передвигающиеся по незнакомой местности, в первую очередь становятся жертвами хищных животных. Аналогичные наблюдения сделаны на пенсильванских полевках и хомячках.
Вот как описывает жизнь домовых мышей в деме известный английский зоолог П. Кроукрофт (1970). «Мышь-самец влачит самое жалкое существование, если только он не принадлежит к «власть имущим». Даже при избытке корма и отсутствии хищников такой самец умудряется выжить только потому, что у него есть товарищи по несчастью и агрессивность доминирующего самца рассредоточивается между ними всеми… Доминирующие самцы не приближаются к кормящим матерям, а они спокойно терпят присутствие не только своих дочерей, но и сыновей — до тех пор, пока те совсем не вырастут. Таким образом, в перенаселенной выгородке в некоторых укрытиях для гнезд сосредоточивалось значительное население, но в основном это были самки и их потомство различного возраста. В эту массу мышей потихоньку втиралось несколько взрослых самцов. Возможно, общий запах гнезда настолько приставал к ним, что маскировал их взрослость. Но как бы то ни было, им удавалось прижиться там, и невольно начинаешь подозревать, что отцы большинства потомства — они, а не доминирующий самец. Хозяева участков были как будто слишком заняты тем, что выгоняли посторонних самцов и пренебрегали своими супружескими обязанностями».
На участие в размножении подчиненных самцов было обращено внимание при исследовании тетеревов (А.М. Чельцов-Бебутов, 1965), кур (Л.З. Кайданов, 1967), макак-резусов (S. Duvall at al., 1976). Но во всех этих случаях вклад таких самцов в оставление потомства был небольшим.
Структурированность популяции связана с регуляцией численности и плотности населения. Рассмотрим механизмы этого процесса на примере мышевидных грызунов, у которых они изучены весьма полно. Накоплено достаточное количество достоверных свидетельств о регулярных циклических колебаниях указанных популяционных параметров, которые не зависят непосредственно от урожайности кормовых растений. А ведь именно этот фактор должен, казалось бы, непосредственно влиять на численность грызунов. Важные наблюдения сделаны, например, Т.В. Кошкиной при учете динамики населения рыжих и красных полевок, а также норвежского лемминга в северной тайге на Кольском полуострове. Приостановка роста численности зверьков наступает обычно задолго до истощения средовых ресурсов. Достижение некоего порога плотности и частоты прямых контактов приводит к изменению физиологического состояния особей. У многих из них развивается состояние психического (эмоциального) стресса (по Г. Селье). Эта «болезнь адаптации» сопровождается характерными признаками — гипогликемией, увеличением надпочечников, гематологическими сдвигами, появлением язв и некрозов. Усиливается агрессивность, возникают аномалии полового цикла. У беременных самок происходит резорбция зародышей. При продолжающемся воздействии стрессорных факторов возрастают нарушения гормонального равновесия и может наступить гибель. Краху популяции способствуют неблагоприятные погодные условия: низкие температуры, дожди, ранние заморозки, поздний неустойчивый снеговой покров, оттепели, гололед и т.д. Но даже в таких условиях часть популяции выживает, и здесь в полной мере проявляется роль дифференциации ее состава.
Территориальные особи и те, которые занимают высокий иерархический ранг во внутрипопуляционных группировках, оказываются относительно устойчивыми к стрессорным воздействиям. Они способны выжить и дать начало следующему поколению. Гибнут прежде всего молодые особи, вынужденные мигрировать, нетерриториальные, а также имеющие низкий зоосоциальный ранг. Эта закономерность подтверждена во многих экспериментах с искусственными группировками домовых мышей, полевок, хомячков, крыс. Таким образом осуществляется авторегуляция плотности населения, связанная со структурированностью популяции. Отбор при этом действует в высшей степени избирательно и надежно. К воспроизведению потомства допускаются только те особи, которые проходят длительную жесткую апробацию на всех стадиях жизненного цикла.
Динамика численности и состава популяционных группировок на основе механизмов авторегуляции выступает как общебиологическое явление и обнаруживается у видов, стоящих на разных ступенях эволюционной лестницы.
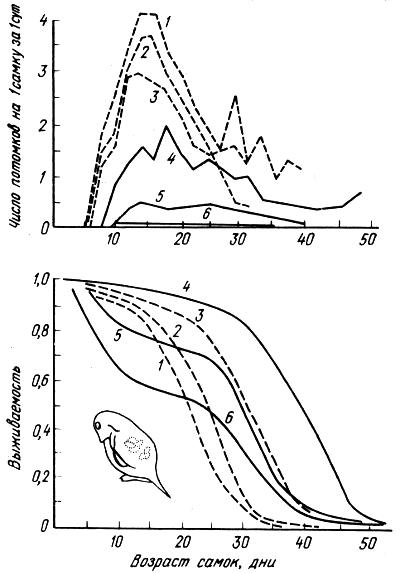
Рис 2.16. Плодовитость и выживаемость самок Daphnia pulex при различных плотностях популяции (по Р. Риклефсу, 1979): число дафний в 1 мл: 1 — одна, 2 — две, 3 — четыре. 4 — восемь, 5 — 16, 6 — 32
На рис. 2.16 показана динамика искусственных популяционных группировок дафний. Пищей дафниям служили одноклеточные зеленые водоросли, которые давались с избытком. Наилучшая выживаемость наблюдалась при плотности 8 особей на 1 мл. При более высоких плотностях индивидуальный рост дафний подавлялся. Численность популяций стабилизировалась при плотности 20 особей на 1 мл среды.
У мучного хруща плотность населения в сосудах с кормом разного объема также устанавливалась на одном и том же уровне. Одним из механизмов авторегуляции служило выделение особями в среду особого вещества, — этилхинона, который ингибирует рост и развитие личинок, а также тормозит размножение имаго. Другой механизм заключался в каннибализме, поедании взрослыми особями откладываемых яиц. Личинки многоножек в крупных скоплениях выделяют газообразный секрет с ощутимым запахом синильной кислоты, тормозящий развитие. На скорость роста личинок ряда видов бесхвостых амфибий влияют продукты метаболизма, которые отрицательно действуют на головастиков, отставших в росте, и одновременно стимулируют развитие тех, которые вырвались вперед. Это явление сочетается с каннибализмом. Каннибализм в качестве механизма, регулирующего плотность населения, отмечен у рыб, например, у гуппи, и даже у высших позвоночных. У зеленых черепах самки при достижении высокой плотности часто уничтожают отложенные кладки яиц. В аналогичной ситуации белые аисты выкидывают из гнезд часть птенцов, обычно более слабых, или просто оставляют их. Сходная картина наблюдается и у других видов птиц.
Вместе с тем мощными и широко распространенными в животном мире механизмами регуляции выживания и размножения являются те, о которых сказано выше, — территориальность и зоосоциальная иерархия. Но и они составляют лишь часть факторов биотической природы наряду с межвидовой конкуренцией, давлением хищников, паразитов, инфекциями и прочими, которые на фоне действия абиотических факторов определяют в конечном счете динамику численности и состава популяций. При этом можно согласиться с Ч. Дарвином, который полагал, что особи того же вида вступают в наиболее сильную во всех отношениях конкуренцию. Через внутривидовые отношения на популяционном уровне осуществляется точная оценка интегративного комплекса признаков, необходимых для воспроизводства потомства.
Структура популяций растений. Специфическое свойство растительных организмов состоит в их прикрепленности к субстрату. Этим определяется пространственная структура популяций растений, отличающаяся от таковой популяций животных. В ее основе лежит система ценопопуляций, или микропопуляций (ценоз от греч. koinos — общий). Ценопопуляциями называют мелкие пространственные группировки растений (совокупности особей одного вида в пределах фитоценоза), до некоторой степени отвечающие понятию дема у животных. Однако сходство здесь весьма отдаленное, поскольку само понятие особь в фитоценологии неоднозначно. В качестве элемента ценопопуляций могут выступать: особь семенного поколения, партикула (особь вегетативного происхождения), часть особи (побег, парциальный куст и пр.), клон (совокупность особей вегетативного происхождения).
Принципиальное значение имеет разнообразие жизненных форм и способов воспроизведения растений. По жизненным формам растения делят на деревья, кустарники, полукустарники, кустарнички, травы — стержнекорневые, короткокорневищные, длиннокорневищные, луковичные и клубневые геофиты; в зависимости от длительности жизни различают одно- и многолетние монокарпии, отмирающие после плодоношения, а также поликарпические многолетники, отмирание которых растягивается на длительный срок. Размножение растений осуществляется половым путем или вегетативным. Возможность вегетативного размножения заложена в самой структуре растительного организма с относительной автономностью отдельных его частей. Оплодотворение может быть перекрестным (аллогамия) или самооплодотворением (автогамия). Широко распространен в растительном мире апомиксис — вторично бесполое размножение. Апомиксис выступает в двух основных формах: в форме партеногенеза, когда начало новому организму дает половая (яйцевая) клетка, и в форме апогамии, при которой новый организм возникает из вегетативной клетки. У апомиктов, цветковых растений, самая распространенная форма — партеногенез, редуцированный (зародыш гаплоидный) и нередуцированный (зародыш диплоидный). Вегетативное размножение и апомиксис ведут к образованию клонов. Лишь при аллогамии в размножении участвуют две родительские особи, в остальных случаях — одна.
Разные способы размножения у одних и тех же видов нередко сочетаются друг с другом. По оценке А. Густафсона, в скандинавских флорах до 80% всех покрытосеменных способны к той или иной форме вегетативного размножения. Многие апомикты воспроизводятся также и половым путем (факультативная агамоспермия). Для гермафродитных покрытосеменных характерен широкий спектр систем скрещиваний — от облигатного переопыления до полной автогамии. Неполной и частичной может быть также самосовместимость.
При изучении структуры популяций растений устанавливают границы ценопопуляций, характер размещения в них растительных элементов, способы их воспроизведения. Растениям свойственно образование скоплений, размеры и плотность которых зависят от дальности распространения семян, роста и развития побегов, степени неоднородности среды, взаимодействия организмов своего и чужих видов в фитоценозах. Важное значение у цветковых аллогамных растительных форм имеет пыльцевой режим — способы и дальность переноса пыльцы.
Осины в природе встречаются, как правило, в виде клонов, образующихся за счет мощных корневых отпрысков от пней. Отдельные клоны осин Populus tremuloides в штате Юта (США) занимают десятки гектаров и насчитывают десятки тысяч деревьев. Их возраст достигает 8000 лет. Многолетний луговой злак Festuca rubra размножается как вегетативно, при помощи корневищ, так и половым путем. Пыльца переносится ветром. Анализ ценопопуляций этого злака в Шотландии показал, что они включают ограниченное число клонов. Их возраст составляет сотни лет. В то же время у другого родственного злака Festuca ovina образуется много небольших по размеру клонов. Здесь соотношение бесполого и полового размножения сдвинуто явно в сторону последнего.
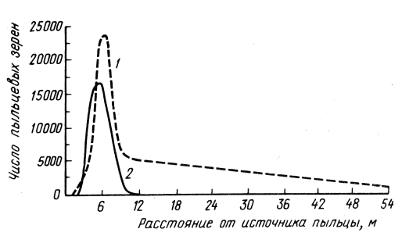
Рис. 2.17. Распространение сосновой пыльцы от родительского дерева (по R.N. Colwell, 1951): 1 — в направлении ветра, 2 — под углом 45° к направлению ветра
Кривые распределения пыльцы от родительских растений до мест падения часто обнаруживают эксцесс и асимметрию (рис. 2.17). Из данного рисунка следует, что большая часть сосновой пыльцы пролетает на ветру от 3 до 10 м, а дальше количество оседающей пыльцы быстро уменьшается. У крымской сосны в горных условиях зона свободного переопыления составляет 300—400 м по вертикали. У переопыляемого пчелами хлопчатника пыльца переносится в основном в радиусе 25 м. У люцерны, переопыляемой разными перепончатокрылыми, дальность разноса основной массы пыльцы составила 50 м, но некоторую ее часть находили на расстоянии километра и даже полутора километров.
Л. Ф. Семериков и Н.В. Глотов провели тщательное исследование пыльцевого режима в популяциях скального дуба на Северо-Западном Кавказе. Сплошные древостои этих деревьев покрывают склоны хребтов на высоте 200—1600 м и отделены друг от друга ущельями, занятыми буком и грабом. Установлено, что расстояние в 70—100 м внутри древостоя является предельным для распространения пыльцы. Пыльца задерживается огромной поверхностью листьев, стволов, ветвей. В буково-грабовых дубняках, где дубы составляют не более 20—30% всего насаждения, возможны случаи практически полной репродуктивной изоляции отдельных деревьев.
В ценопопуляциях многих видов свободно скрещивающиеся группы включают всего несколько десятков или сотен растений.
Плотность растений в ценопопуляциях влияет на интенсивность их роста и соотношение органов. У многолетников при загущении интенсифицируется корнеобразование и падает биомасса, приходящаяся на корневища или столоны, уменьшается вероятность образования генеративных побегов. Для многих однолетников выявлено отрицательное влияние возрастания плотности на семенную продуктивность и одновременно положительное влияние на вероятность перекрестного опыления. В соответствии с правилами В.Н. Сукачева при увеличении плотности у однолетников развитие ускоряется, у многолетников замедляется. Не одинаково протекают у тех и других процессы самоизреживания. Оставшиеся в живых мелкие однолетники не способны компенсировать свой рост даже в самых благоприятных условиях, тогда как многолетники такую компенсацию осуществляют. Оптимизация популяционной структуры у разных видов достигается по-разному, при этом под контролем отбора регулируются плотность, биомасса и возрастной состав растений.
В этой главе были рассмотрены дифференциация популяционной структуры и влияние на нее абиотических и биотических факторов среды.
В популяциях позвоночных эколого-физиологическая дифференциация особей определяется их распределением в пространстве, владением участками территории (акватории) или же местом, которое они занимают во внутривидовых группировках. Особи, остающиеся без индивидуальных участков или оказывающиеся на низких ступенях зоосоциальной иерархии, полностью или частично устраняются от оставления потомства. Они нередко отличаются измененным гормональным статусом, недоразвитием репродуктивной системы, характерными особенностями поведения. К ним относятся большинство молодых половозрелых животных, а также значительная часть животных старших возрастов. Внутрипопуляционный резерв особей, устраняемых от размножения, оказывается очень большим, особенно при неблагоприятных условиях. К воспроизведению потомства допускаются лишь немногие индивиды, которые прошли жесточайшую внутривидовую и межвидовую конкуренцию и вышли победителями. Оценить напряженность и эффективность отбора можно только при раскрытии организации группировок внутри популяций, закономерностей их поддержания и преобразования.
При жесткой зоосоциальной иерархии эффективный размер популяции, о котором будет сказано в гл. 4, в значительной степени определяется неравномерным участием в размножении особей женского и мужского пола, устранением основной массы самцов. Степень замкнутости внутрипопуляционных группировок, как и система размножения в них, непосредственно отражается на коэффициентах инбридинга. Интенсивность вытеснения мигрантов и возможность их внедрения в чужие группировки влияют на «поток генов», частоту обмена генетической информацией.
Структурированность популяций растений и степень замкнутости ценопопуляций, способы их воспроизведения также тесно связаны с направлением отбора и имеют важнейшие генетические последствия.
