2.4. ПОПУЛЯЦИЯ В СИСТЕМЕ БИОГЕОЦЕНОЗА
Биогеоценозами называют сложившиеся естественно-историческим путем элементарные подразделения биосферы Земли. Они включают почву, бактерии, грибы, растения, животных, которые все вместе составляют экосистему и существуют в конкретных климатических условиях. Биотические компоненты экосистемы образуют сообщество и состоят из автотрофов и гетеротрофов. Автотрофы — зеленые растения — превращают солнечную энергию в химическую. Гетеротрофы, или консументы, — животные, поедающие растения или других животных. К гетеротрофам относят также деструкторы — бактерии, грибы, некоторые растения. Они расщепляют органические вещества на более простые компоненты и замыкают тем самым круговорот веществ в биогеоценозе.
Своеобразные оазисы жизни найдены за последние четверть века на дне Тихого и Атлантического океанов. Основу их существования составляет поток органического вещества не с поверхности, а придонный бактериальный хемосинтез — использование бактериями энергии, получаемой при окислении органических соединений, выносимых гидротермальной циркуляцией.
Биопродукция таких сообществ достигает 40—60 кг на 1 м2. Они включают многощетинковых червей, брюхоногих и двустворчатых моллюсков, веслоногих рачков, представителей других групп животных.
Экологические процессы в популяциях, такие, как размножение, рост, образование биомассы, определяются многими факторами абиотической и биотической природы. Особое значение имеют лимитирующие факторы: температура, влажность, нехватка пищи, мест для размножения и др. Степень видовой специализации, очевидно, прямым образом влияет на возможности приспособления к меняющимся условиям среды. Приспособление, или адаптация, может быть определено как соответствие отдельных особей или групп особей окружающей среде. При этом речь должна идти об адаптивных комплексах признаков, имеющих наследственную обусловленность и реализующихся в пределах норм реагирования в процессе индивидуального развития.
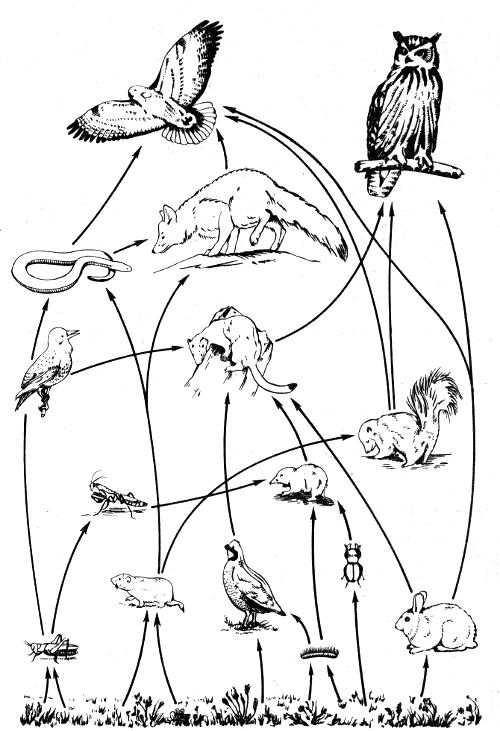
Рис. 2.8. Пищевые связи в упрощенной трофической системе (по Р. Риклефсу, 1979)
Основу отношений между популяциями разных видов в экосистемах составляют пищевые связи. Они исключительно сложны и многообразны. На рис. 2.8 указаны некоторые пищевые связи в относительно простой трофической системе. Здесь представлены растительноядные и плотоядные животные, хищники, которые убивают и пожирают своих жертв. Паразиты, в отличие от хищников, кормятся на живой жертве, или хозяине, а сапрофиты питаются мертвечиной. Поглощение пищи, т.е. хищничество в самом широком смысле, представляет собой главный фактор передвижения материалов и энергии в экосистеме. Поэтому изучение хищничества дает ключ для понимания процессов, определяющих функционирование сообществ.
Теоретические основы динамики популяций хищника и жертвы были заложены в 20-х годах американским ученым А. Лоткой и итальянским ученым В. Вольтеррой. Они предложили систему кинетических дифференциальных уравнений, с помощью которых показали возможность возникновения циклических незатухающих колебаний плотности популяций хищника и жертвы.
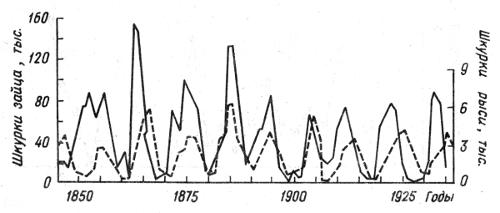
Рис. 2.9. Связь колебаний численности американского зайца-беляка Lepus americanus и канадской рыси Lynx canadensis (по D. MacLulich, 1937): сплошной лившей обозначены колебания численности популяции зайца-беляка, прерывистой — популяции рыси
На рис. 2.9 представлены хорошо документированные за 100-летний период данные сопряженных изменений плотности популяций зайца-беляка и рыси в районе Гудзонова залива в Канаде. Рис. 2.10 отражает динамику трех видов хищников — рыси, лисицы и волка — и зайца-беляка в центральных районах европейской части России за четверть века. Отчетливо виден циклический, сопряженный характер колебаний численности жертвы и хищников. Плотность популяций в отдельные периоды меняется в десятки раз. Механизм зависимости в этих и других случаях на первый взгляд достаточно прост. Хищники, увеличиваясь в числе, быстро сокращают численность жертв и тем самым подрывают свою кормовую базу. В результате большая часть хищников гибнет от голода. При ослаблении с их стороны прессинга на популяцию жертвы последняя интенсивно размножается и весь цикл начинается сначала. В действительности механизм описываемых явлений более сложный. Об этом еще будет сказано в данной главе. К тому же найдены зависимости иного рода. Так, обыкновенная неясыть, несмотря на значительные колебания численности ее жертв, мелких млекопитающих, длительное время сохраняет Свою численность на постоянном уровне (рис. 2.11).
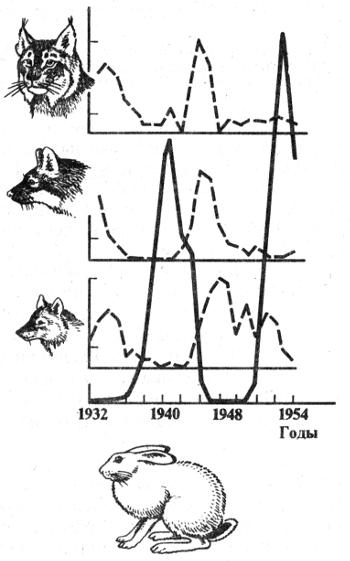
Рис. 2.10. Характер колебаний численности рыси Felis lynx, лисицы Vulpes vulpes, волка Canis lupus и зайца-беляка Lepus timidus на севере европейской части России (по А.В. Яблокову, А.Г. Юсуфову, 1989)
Если хищничество направлено на одностороннюю в общем выгоду одной из взаимодействующих популяций — хищника — и в меньшей степени жертвы, то конкуренция представляет собой симметричный процесс. Межвидовая конкуренция возникает тогда, когда две (или более) популяции близких видов используют одни и те же ресурсы, имеющиеся в недостатке. Важнейший вклад для понимания происходящих при этом событий внесли лабораторные эксперименты Г.Ф. Гаузе, выполненные в начале 30-х годов в Московском университете.
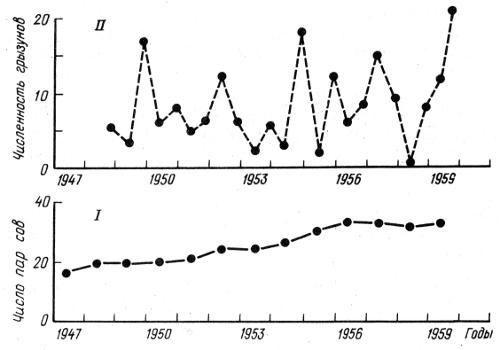
Рис. 2.11. Численность в системе хищник—жертва обыкновенной неясыти Strix aluco (I) и мелких млекопитающих — лесных мышей и рыжих полёвок (II) (по Н. Southern, 1970)
Гаузе экспериментировал с простейшими — ресничными инфузориями Paramecium aurelia, P. caudatum и Р. bursaria. Популяции всех трех видов успешно поддерживались в монокультурах, достигая предельных плотностей. Пищей инфузориям служили дрожжевые или бактериальные клетки. В смешанных культурах P. aurelia чаще всего вытесняла P. caudatum, вплоть до полного вымирания последней (рис. 2.12). Совсем иначе складывались конкурентные отношения между P. aurelia и P. bursaria. Искусственные популяции этих двух видов могли неопределенно долго сосуществовать друг с другом, хотя плотности их в условиях конкуренции были ниже, чем в монокультурах. Возможность сосуществования определили, как было выяснено, различия в пищевой специализации. Особи Р. bursaria селились в основном на дне пробирок и питались оседающими дрожжевыми клетками. Особи P. aurelia, напротив, держались в верхней части пробирок и питались преимущественно бактериями. Исходя из полученных результатов Гаузе определил экологическую нишу как место, которое вид занимает в экосистеме. В каждом сообществе виды выполняют свои функциональные роли. Крупным обобщением явился принцип конкурентного исключения, или закон Гаузе, который гласит: два вида, занимающие общую экологическую нишу, не могут длительное время существовать совместно.
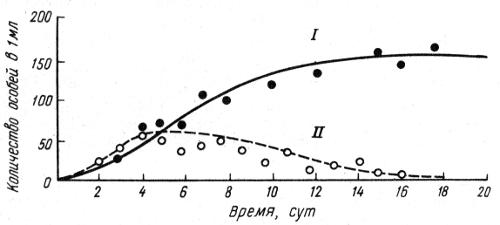
Рис. 2.12. Динамика конкурентных отношений двух видов инфузорий — Paramecium aurelia (I) и P. caudatum (II) в экспериментальной культуре (по Г.Ф. Гаузе, 1934)
Принцип конкурентного исключения получил многочисленные подтверждения при исследовании природных и искусственных сообществ. Рассмотрим несколько примеров. При изучении использования пространства в популяциях пяти симпатрических видов американских славок установлено, что особи каждого вида отличаются своим специфическим характером использования разных частей деревьев хвойного леса (рис. 2.13). Они также неодинаковыми способами ведут поиск насекомых среди веток и хвои.
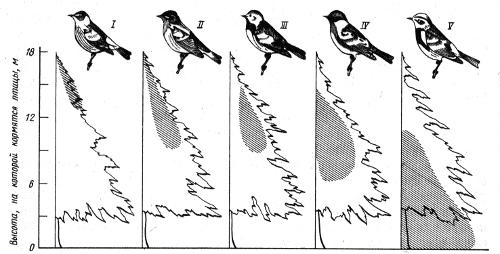
Рис. 2.13. Локализация мест кормежки пяти видов американских славок Dendroica в еловых лесах штата Мэн (по R.H. MacArthur, 1958): I — D. tigrina. II — D. fusca. III — D. virens. IV — D. castanea, V — D. coronata
Различными рационами характеризуются симпатрические морские моллюски рода Conus, ящерицы рода Ctenotus, родственные симпатрические виды многих птиц и млекопитающих. При совместном выращивании овса двух близкородственных видов, Avena barbata и A. fatua, была установлена лучшая конкурентоспособность второго вида, растения которого в посевах различной плотности продуцируют больше семян. Мучные хрущаки Tribolium castaneum и Т. confusum обнаруживают неодинаковую конкурентоспособность в зависимости от условий среды. При невысокой температуре и низкой влажности Т. castaneum вытесняет Т. confusum, а в теплой и влажной среде результат оказывается прямо противоположным.
! Следствия эволюции системы хищник — жертва заключаются в увеличении эффективности поиска и ловли жертвы в популяциях хищника и в совершенствовании избегания в популяциях жертвы. Хищники выступают в качестве регулятора плотности популяции жертвы, удерживая ее на уровне, оптимальном для экосистемы. Эволюционные последствия конкуренции состоят в оптимизации использования ресурсов среды у близких видов за счет их дивергенции по пищевым и прочим потребностям. При этом также достигается совершенствование экосистем как целостных образований. Ведущим фактором эволюции экосистем служит естественный отбор, который контролирует характер межвидовых отношений прежде всего через пищевые связи.
