20.7. Современный сальтационизм
Существуют представления о сальтационном, внезапном образовании новых видов, вне действия отбора, путем крупных скачков. Эта идея своими корнями восходит к глубокой древности. В XX в. эти идеи старались обосновать, обращая внимание на наличие «организационных признаков», выходящих якобы за рамки обычных приспособлений (А. Келликер, Э. Зюсс, О. Шиндевольф), или внезапное появление видов путем крупных мутаций (С.И. Коржинский, Г. де Фриз, В. Бэтсон, Р. Гольдшмидт).
Такие подходы (сальтационизм) оспаривают дарвиновский тезис о постепенном ходе эволюции (градуализм). Заметим, что сам Ч. Дарвин, хотя вслед за Лейбницем отмечал отсутствие в природе скачков («Natura non facit saltum»), не отрицал возможность возникновения эволюционных новшеств «по законам роста и корреляций».
В последнее время идея о скачкообразном характере возникновения видов вновь находит сторонников как среди генетиков, так и со стороны палеонтологов. Учитывая, что среди них находятся исследователи, которых трудно заподозрить в профанации, остановимся на этих взглядах несколько подробнее.
Для многих групп организмов известны так называемые макромутации — мутации, результатом которых является появление у особи признака, характерного для других крупных таксономических групп. Например, мутация tetraptera ведет к появлению у дрозофилы и домовой мухи второй пары крыльев — признака другого отряда насекомых. В результате мутации aristopedia у насекомых вместо антенны возникает нога. Такие системные мутации заметно меняют ход онтогенеза. Мутации hairless и nude ведут к появлению голых (безволосых) мышей, кошек, собак, овец. Мутация мопсовидности ведет к редукции межчелюстных костей и, как следствие, сокращению размеров лицевой части черепа у человека, собак, кошек, свиней. Подобные факты привели Р. Гольдшмидта в 40-е годы XX в. к представлению, что эволюция может происходить за счет фиксации резких перестроек морфогенеза, вызванных единичными системными мутациями — через возникновение «многообещающих монстров». В разной форме эту идею поддержали такие видные эволюционисты, как К. Уоддингтон, С. Гоулд и Н.Н. Воронцов. М. Уайт, Н.Н. Воронцов и Р. Маттей развили концепцию внезапного формо(видо)образования на основе хромосомных перестроек на большом материале по кариосистематике грызунов и других млекопитающих, напоминая о гибридогенном происхождении сливы (терн х алыча), экспериментальном синтезе редько-капусты (Raphanobrassica), о горизонтальном переносе мобильных диспергированных генов проявляется один из механизмов.
В 60-е годы XX в. было показано широкое распространение дупликации генов (удвоения участков ДНК) при образовании полипептидных цепей гемоглобина и других сложных белков (С. Оно). Фермент карбоангидраза у плацентарных и сумчатых млекопитающих образован двумя полипептидными цепями, а у беспозвоночных, рыб и птиц — одной. Эта дупликация могла произойти при возникновении млекопитающих около 200 млн лет назад (см. рис. 17.2).
Нам представляется, что все эти факты и теоретические предположения (в числе последних — горизонтальный перенос) не могут рассматриваться как противоречащие дарвиновской концепции постепенного видообразования. Одна мутация безволосости у предков современного моржа не могла привести к возникновению безволосости как видового признака без многочисленных дополнительных адаптивных изменений в строении кожи, подкожной жировой клетчатки, механизма кровоснабжения и терморегуляции и пр. Возникновение (и распространение в пределах вида) этого комплекса адаптаций должно было происходить на протяжении множества поколений. Любой «многообещающий монстр», возникший в результате одиночной мутации, должен был пройти все обычные стадии микроэволюции:
— должна была возникнуть не одна, а хотя бы небольшая, но группа измененных особей;
— должна была возникнуть генетическая изоляция такой группы от исходной формы;
— такая форма видового ранга (см. 13.4) должна была пройти «шлифовку» естественным отбором на протяжении многих поколений, прежде чем образовать собственную экологическую нишу и стать видом (см. гл. 11 и 12).
Убыстрение эволюции, несомненно, происходящее после приобретения каких-то принципиальных приспособлений (например, ароморфозов) и обеспечивающее выход группы в новую адаптивную зону (см. анагенез, разделы 15.3, 15.5), в эволюционном масштабе может внешне выглядеть как скачок, сальтация, не являясь таковым на самом деле.
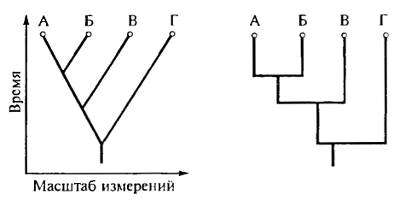
Рис. 20.4. Сопоставление постепенной («дарвиновской») и прерывистой (пунктуалистской) эволюции групп. А—Г — виды. Уровень различий между видами и время дивергенции отдельных филумов полностью совпадают в обоих случаях (ориг.)
Второй фронт современного сальтационизма — гипотеза прерывистого равновесия или пунктуализма (С. Гоулд, Ф. Стенли, Н. Элдридж). Суть пунктуализма состоит в допущении, что эволюция идет только скачками, в промежутки между которыми наблюдается длительный застой — стазис (рис. 20.4). При изучении ископаемых остатков пресноводных моллюсков в отложениях африканского озера Туркана, а также некоторых рептилий и млекопитающих не удалось обнаружить переходных форм между видами, характерными для разных непрерывных горизонтов отложений. Периоды же стазиса для некоторых видов моллюсков, по их подсчетам, продолжались 105—107 поколений. Затем виды моллюсков за сравнительно короткое время (103—104 поколений) якобы обнаруживали изменчивость и давали начало дочерним видам. По мнению пунктуалистов, таким путем образованы примерно 95% видов. Это должно было произойти в результате преимущественного преобразования периферических изолятов в пределах единой популяции и без участия отбора (вымирание предкового вида допускается без конкуренции, новый и предковый виды не отличаются по адаптивности).
При объяснении вымирания некоторых крупных рептилий сальтационистские тенденции проявляются в поиске причин глобальных катастроф космического характера — падение метеоритов, геомагнитные инверсии, резкое повышение радиационного фона Земли из-за изменения солнечной активности и т. д. Сальтационизм проявился и в молекулярной биологии в стремлениях объяснить возможности быстрого возникновения видов и более крупных таксонов на основе случайного заноса целых блоков генов (мобильных генетических элементов) от низших организмов к высшим или от одних групп организмов к другим с помощью фагов и вирусов (горизонтальный перенос генов). При таком переносе предполагают возникновение крупных изменений вне действия естественного отбора. В этом же ключе объясняется даже быстрое возникновение Человека разумного как результат резких мутаций, которые могли иметь место в районах повышенного радиационного фона в Африке.
Было бы неправильно огульно отвергать возможности эволюционных изменений на основе сальтаций. Факты говорят о том, что они вполне вероятны и реальны. Так, на примере растений допускается возможность внезапного (быстрого) эволюционного преобразования спорангия (путем агрегации и срастания спороносных частей водорослей), семяпочек (путем таких же изменений макроспорангиев), преобразования четырехчленного цветка из пятичленного, однодольных из двудольных и т. д. (А.Л. Тахтаджян).
Сальтационные изменения и их эволюционная фиксация характерны и для животных: перенос периферических сосудов или нервов с одного ствола на другой (например, перенос сонных артерий у птиц на правую дугу аорты, а у млекопитающих — на левую, уменьшение или увеличение числа шейных или туловищных позвонков и т. д.). Однако возможность возникновения признаков, видов и даже других таксономических единиц на основе каких-то резких уклонений не дает оснований для принятия гипотез пунктуализма и тем более сальтационизма.
Существуют объективные трудности в анализе процесса изменчивости на ископаемом материале, во-первых, ввиду его редкости, что не позволяет исключить возможных переходов между видами (рис. 20.5), во-вторых, из-за возможных смещений места захоронения организмов после их гибели (о переносе на большие расстояния водой и смещении осадочных пластов писал еще Ч. Дарвин); в-третьих, есть данные (в том числе и для ископаемых моллюсков оз. Туркана) об одновременном возникновении эволюционных изменений (в результате оттеснения предкового вида на прилегающие территории); в-четвертых, связь между скоростью морфологической эволюции и процессами видообразования, как показывают многочисленные современные данные, отсутствует.
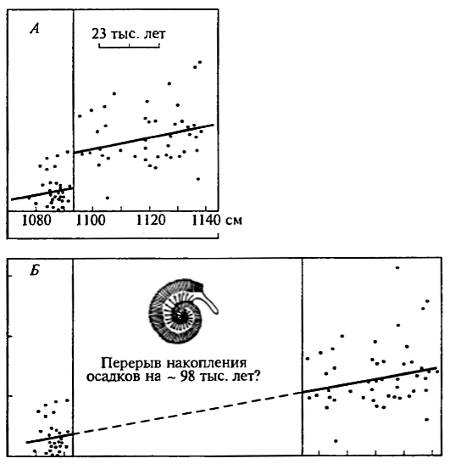
Рис. 20.5. Пример ненадежности сальтационной интерпретации палеонтологических данных (по Д. Раупу и Д. Стенли, 1974)
В верхнеюрских оксфордских глинах Питерсборо (Англия) обнаружены ископаемые аммониты рода Cosmoceras. Эти отложения возникли, по расчетам, со скоростью 1 м за 115 тыс. лет. Важной характеристикой аммонитов является отношение числа ребер к числу краевых шипов на раковине: А — реальное стратиграфическое положение каждого из аммонитов, обнаруженных в слое 1080—1140 см. Линия обозначает среднеарифметическое значение отношения числа ребер к числу шипов. В слое 1093,5 см соседние значения резко смещены, что дает основание для предположения о сальтации признака; Б — части того же графика разнесены таким образом, чтобы восстановить последовательность постепенного изменения морфологии раковин. Альтернативной сальтационному оказывается вполне реальное предположение, что на протяжении 98 тыс. лет осадки в данном месте почему-то не накапливались
Американский палеобилог Дж. Г. Симпсон, обсуждая роль горизонтального переноса генов в эволюции заметил, что такое явление «не остается за границами этой (синтетической — авт.) теории, а раздвигает их шире» (В. Грант. Эволюция организмов. М., 1980. С. 12). То же самое можно сказать и о других фактах, лежащих в основе разного рода сальтационных взглядов. Пока не найдено ни одного доказательства, что исходный механизм эволюции в природе — возникновение видов — носит не приспособительный характер, «обходится» без естественного отбора и возникновения адаптаций.
Выше были рассмотрены лишь некоторые из современных проблем, стоящих перед эволюционным учением. Мы не затронули важной проблемы проявления эволюционных принципов на разных уровнях организации живого; не сказали о важности выяснения пути от гена к фену (создание пока отсутствующей теории индивидуального развития, окажет мощное влияние и на развитие эволюционной теории).
Может быть, когда будут выяснены основные закономерности эволюции биогеоценозов, создана теория индивидуального развития и обнаружены иные пути развития живого во Вселенной, тогда современное эволюционное учение, основанное на теории естественного отбора, окажется лишь составной частью какой-то общей теории развития органического мира. Пока же эволюционная теория с каждым новым принципиальным открытием в биологии укрепляется. Так было после открытия основных генетических закономерностей в начале XX в., так было в середине XX в., после открытия генетического кода и успехов в молекулярной биологии, так, мы уверены, будет и в дальнейшем.
