16.4. Темпы эволюции органов и функций
В предыдущей главе подчеркивалось, что темпы эволюции можно рассматривать как скорость эволюции отдельных форм в целом (видов, родов, семейств и т. д.) и как скорость эволюции отдельных признаков и структур. Скорость эволюции отдельных признаков и органов иногда может быть практически более важной характеристикой. Например, важно знать, как быстро у того или иного вида может возникнуть невосприимчивость (резистентность) к применяемым химическим препаратам: к инсектицидам — у насекомых, к дефолиантам — у растений, к лекарственным препаратам — у человека и т. д.
Скорость эволюции отдельных органов, естественно, тесно связана со скоростью эволюции вида в целом; возникновение или значительное изменение органа всегда является видовым признаком и обычно связано с процессом видообразования. Поэтому скорость эволюции органов практически должна быть близка к скоростям образования новых видов (т. е. должна исчисляться сотнями и тысячами поколений). По-иному обстоит дело со скоростью образования отдельных новых признаков в популяциях или их группах. На протяжении жизни всего лишь десятка поколений отдельные популяции березовых пядениц (Biston betularia) приобрели признак меланизма, с такой же скоростью распространяются внутри вида многие другие признаки: нечувствительность к варфарину у крыс (см. 10.4), устойчивость к пестицидам у ряда видов насекомых и др. (см. гл. 21). Скорость эволюции любого признака или группы признаков, как показывает палеонтологический материал, может значительно изменяться на разных этапах эволюции всей группы, и при этом темпы эволюции признаков могут быть как сходными, так и различными (рис. 16.3). Интересен вопрос относительно скорости эволюции количественных признаков (например, размер тела и его частей). Для измерения скорости эволюции таких признаков предложен показатель, названный «дарвином» (Дж. Холдейн); 1 дарвин соответствует изменению среднего значения (уменьшению или увеличению) данного признака на 1% за тысячу лет. Этот условный показатель учитывает не число поколений (более фундаментальную эволюционную характеристику), а астрономическое время.
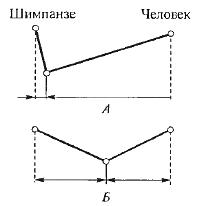
Рис. 16.3. Скорость эволюции разных признаков в предковых линиях шимпанзе (Pan troglodyres) и человека (Homo sapiens): А — масштаб изменений внешнего облика; ? — масштаб изменений белков. Скорость изменения внешнего вида много выше у человека, скорость изменения белков одинакова (из В. Гранта, 1980)
Однако и он дает возможность для некоторых сопоставлений. Так, темп эволюции многих признаков, касающихся размеров тела и скелета, составлял для млекопитающих от десятых долей до нескольких десятков Дарвинов (рис. 16.3, 16.4), близкие величины найдены и для моллюсков.
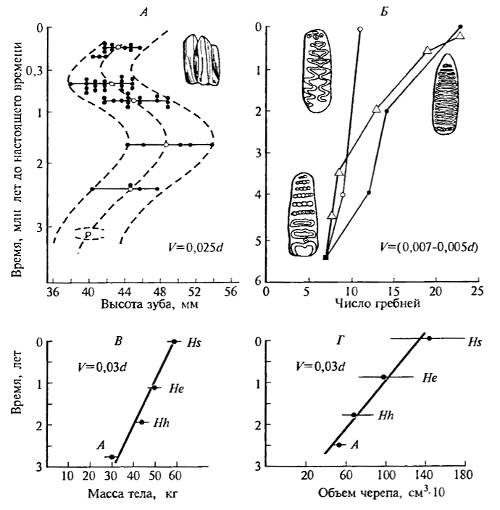
Рис. 16.4. Темпы эволюции некоторых органов и структур: А — величина коренного зуба зубров рода Bison; Б — число гребней коренного зуба в трех линиях слонов: индийского Elephas maximus (точки), африканского Loxodonta africana (кружки) и мамонта Mammuthus primigenius (треугольники); в — средняя масса тела в роде Homo; Г — объем мозговой коробки в роде Homo (A — Australopithecus. Hh — Н. habilis. He — Н. erectus, Hs — Н. sapiens) (по данным разных авторов)
Скорость эволюции отдельных признаков в популяциях, так же как и скорость эволюции целых структур и органов, зависит от многих факторов: численности особей в популяциях, числа популяций внутри вида, продолжительности жизни поколений (рис. 16.5). Однако скорость эволюции слонов, отличающихся не особенно быстрой сменой поколений, как показывают палеонтологические данные, по многим признакам может быть гораздо выше, чем скорость эволюции дрозофилы, имеющей ряд поколений в течение года. Можно сказать, что любые самые сложные факторы первично окажут свое воздействие на скорость изменения популяции и вида посредством изменения давления элементарных эволюционных факторов: мутационного процесса, волн жизни, изоляции и, главное, естественного отбора.
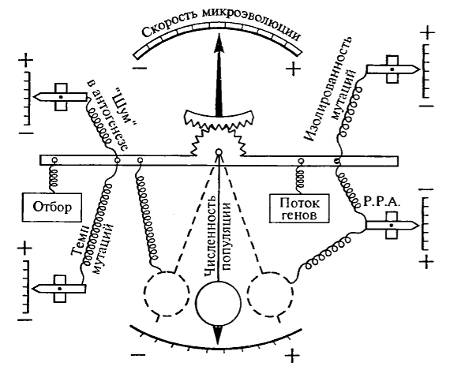
Рис. 16.5. Схема влияния на скорость эволюции различных микроэволюционных факторов. На этой механической модели (предложенной Д. Шпирлихом, 1973) видно разнонаправленное взаимодействие эволюционных факторов. Р.П.А — радиус репродуктивной активности особи (из А.В. Яблокова, 1987)
Мультифункциональность любого органа и способность количественного изменения функции — основы эволюционных преобразований органов.
В эволюции отдельных органов и функций разные принципы морфофизиологических преобразований взаимно переплетаются и выделить их в чистом виде трудно. При этом очевидна связь процессов преобразования функций с процессами преобразования органов: в основе всех преобразований органов в эволюции лежат процессы смены функций.
Процессы макроэволюции, выражающиеся в перестройках органов, тесно и неразрывно связаны с процессами микроэволюции и в конечном счете определяются изменениями популяций и видов. Существо этих процессов всегда состоит в возникновении и изменении адаптаций — адаптациогенезе.
