Географические различия между видами основаны на относительной самостоятельности ареала каждого вида. Несомненно, что размер ареала, его форма, расположение в биосфере — важные видовые признаки. Для каждого вида свойственны свои границы пригодных для жизни условий, своя история возникновения (определяющая в значительной степени очертания и размер ареала), свои специфические взаимодействия с видами-конкурентами и т. д. Поэтому любая характеристика вида будет неполной без указания на специфические особенности его ареала.
Однако существуют причины, по которым географический критерий не может считаться достаточно универсальным для характеристики видов в природе. Во-первых, существует множество видов с практически совпадающими ареалами. Например, ареал кедрового стланика (Pinus pumila) совпадает с ареалами березы Миддендорфа (Betula middendorfii), даурской лиственницы (Larix daurica) и душистого тополя (Populus suaveolens). Эти виды тесно связаны биогеоценотически, входят в состав одних и тех же растительных сообществ, и их историческое развитие шло, видимо, параллельно, отражая общность исторических условий формирования определенных природных зон. Во-вторых, для видов-космополитов, ареал которых охватывает огромные пространства в биосфере Земли (в океане, например, дельфин косатка — Orcinus orca; на суше — некоторые виды рода дрозофил), характеристика ареала как видового признака также теряет смысл. В-третьих, определенность ареала отсутствует у некоторых быстрорасселяющихся видов, поскольку очертания ареала у них изменяются очень быстро и непредсказуемо (в последние столетия часто в связи с антропогенными изменениями ландшафтов или вселениями, как, например, для домовой мухи, городского воробья, многих рудеральных растений и т. д.).
Итак, и географический критерий не может служить каким-либо универсальным «видовым» признаком.
Генетическое единство — главный критерий вида. Целостность вида. Возникает вопрос: а есть ли такой критерий? Есть, и он характеризует вид и как всеобщую форму существования жизни на нашей планете, и как качественный этап эволюционного процесса. Основной критерий вида заключается в его генетическом единстве. Единство вида как динамической системы в эволюции основано на возможности нивелировок различий, возникающих в популяциях путем скрещивания. Как бы ни были изолированы отдельные популяции и подвиды, поток генетической информации между ними всегда существует. Даже единичные миграции отдельных особей из одной популяции в другую, из зоны обитания одного подвида в зону обитания другого, повторяясь на протяжении сотен и тысяч поколений, создают поток генов. Это обеспечивает интеграцию относительно изолированных генофондов отдельных популяций (рис. 12.1).
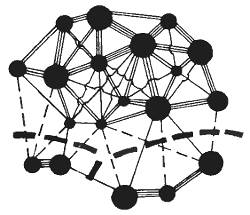
Рис. 12.1. Схема интеграции генофондов отдельных популяций в единый генофонд вида. Кружки — популяции, линии — потоки генов между ними, пунктир — возникающая изоляция частей видового генофонда (ориг. рис.)
Конечно, при отсутствии обмена гамет (у агамных, облигатно-партеногенетических и самооплодотворяющихся форм) не возникает такого генетического единства особей, что создает объективные трудности при разграничении видов.
Единство вида не нарушается даже и в том случае, если иногда по каким-то причинам границы между соседними видами оказываются нечеткими. Сейчас в разных группах таких случаев вскрывается все больше. Это связано с тем, что особи близких видов в определенных условиях иногда могут успешно скрещиваться друг с другом и давать плодовитое потомство в некоторых комбинациях. В таких случаях оказывается, что, за исключением некой гибридной зоны, между такими видами нарушения строения обоих видов как самостоятельных генетических систем не происходит: виды оказываются не генетически закрытыми системами, как представлялось еще в середине XX в., а генетически устойчивыми системами.
Единство и эволюционная «судьба» вида определяют его специфическое место в биогеоценозах. В природе нет двух видов с одинаковым набором адаптаций: это разнообразие адаптаций делает каждый вид уникальным и определяет возможность формирования каждым видом своей экологической ниши как суммы экологических ниш отдельных, составляющих вид популяций.
