Сложные адаптации. В развитии эволюционного учения особое значение имело материалистическое объяснение возникновения некоторых чрезвычайно сложных адаптаций путем накопления мелких наследственных уклонений. Среди таких адаптаций ниже рассматриваются способность к насекомоядности у растений, развитие глаза как органа зрения, возникновение взаимных приспособлений у насекомых и цветковых растений.
Насекомоядность и способность к движению у растений. Хищничество для таких автотрофных организмов, как растения, является исключением, но тем не менее даже среди цветковых встречаются сотни видов насекомоядных растений.
Росянка (Drosera rotundifolia) имеет ряд совершенных и эффективных приспособлений для ловли и переваривания насекомых. Листья у нее покрыты чувствительными волосками, у основания которых расположены секреторные клетки, выделяющие липкую и ароматную жидкость. Стоит насекомому сесть на листовую поверхность, как оно прилипает. Пытаясь вырваться, насекомое бьется, вызывает раздражение чувствительных волосков: раздражение передается по пластинке и вызывает ее свертывание (рис. 11.7). Вслед за этим шаровидные кончики волосков на верхней стороне пластинки выделяют жидкость, богатую ферментом пепсином, под влиянием которого насекомое переваривается и растворенные вещества всасываются листовой пластинкой, затем листовая пластинка расправляется и готова вновь схватить насекомых.
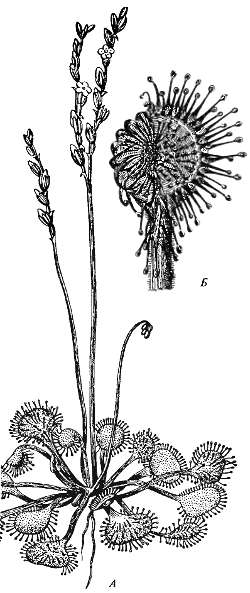
Рис. 11.7. Пример насекомоядного растения — росянка круглолистная (Drosera rotundifolia). A — общий вид растения и Б — ловчий лист, на который был положен небольшой кусочек мяса (ориг. рис. Ч. Дарвина)
У других насекомоядных растений для ловли насекомых есть специальные органы — «кувшины» и «капканы». Так, венерина мухоловка (Dionaea muscipula) ловит мух движением половинок листа. При этом зазубренные концы правой и левой половинок пластинки заходят друг за друга, образуя своеобразный капкан.
Развитие насекомоядности у растений первоначально было связано с поглощением продуктов распада мертвых организмов, случайно попавших на поверхность какого-либо органа. Разложение мертвых организмов могло происходить под влиянием бактерий или гидролитических ферментов растения. В дальнейшем эти возможности растений были не только усилены, но и дополнены ловлей живых насекомых, привлекаемых ароматом выделений или окраской растения. Отбор на насекомоядность у растений был связан с развитием и усовершенствованием способов ловли насекомых, т. е. усовершенствованием ловчего аппарата и ферментов.
Исследования показывают, что ферментативные системы, обеспечивающие переваривание животной пищи растениями, подвергаются типичным мутационным изменениям. На этой основе и должен был действовать естественный отбор, в определенных условиях приведший к возникновению и развитию насекомоядности. Несомненно, ловчий аппарат первых насекомоядных в далеком прошлом был примитивным, совершенство и разнообразие ловчих органов растений возникло позже — в процессе естественного отбора более удачных вариантов.
Не менее интересны приспособления растений к разнообразным движениям (тропизмы, лазание и т. п.), впервые с эволюционных позиций проанализированные в специальных работах Ч. Дарвина. Интерес Ч. Дарвина к этому вопросу не случаен. Его критики полагали, что принципом отбора нельзя объяснить, в частности, своеобразные особенности лазающих растений. Подробно рассмотрев формы движения растений, Дарвин пришел к выводу, что способность к движениям, присущая многим растениям, у части видов в ходе эволюции была усилена, если подхватывалась естественным отбором. Предпосылки для этого в виде широкой изменчивости гормонов, определяющих темпы ростовых процессов, существуют внутри каждого вида растений (такие мутации получены и экспериментально). Каждое растение обладает способностью к ростовым движениям в той или иной степени. Эта способность может быть усилена в результате естественного отбора мутаций, связанных с морфологическими, биохимическими и физиологическими признаками.
Строение органа зрения. 140 лет назад возник спор: можно ли объяснить возникновение такого сложного органа, как глаз, накоплением мелких случайных уклонений? Находятся и сегодня биологи, которые заявляют: «Достаточен один вид глаза, чтобы понять невероятность его развития с помощью естественного отбора». Анализ этого возражения против теории естественного отбора был в наиболее полном виде сделан еще самим Ч. Дарвином, рассматривавшим критику его взглядов Ст. Майвартом. Анализируя возражения Майварта, Ч. Дарвин показал, что глаз как орган зрения возник не сразу, его развитие проходило путем постепенного усовершенствования (рис. 11.8). У части одноклеточных организмов нет восприятия света, у других — на переднем конце тела появляется светочувствительное пятно, насыщенное пигментом.
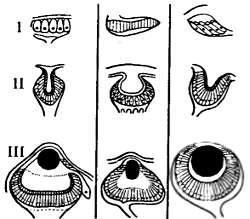
Рис. 11.8. Последовательные стадии усложнения глаза у кольчатых червей (I), членистоногих (II) и моллюсков (III). Развитие органа зрения от группы светочувствительных клеток до совершенных структур, концентрирующих свет и фокусирующих изображение, в ходе эволюции происходило в разных группах животных (по В.Н. Беклемишеву, 1952)
Эволюция многоклеточной организации связана с усложнением аппарата восприятия света. Так, уже у планарий (плоские черви) появляются примитивные «глазки» — чашевидные углубления, заполненные пигментом родопсином.
У более высоко организованных существ строение глаза усложняется. У членистоногих появляются глаза фасеточного типа. Отдельно взятая фасетка, хотя и снабжена группой светочувствительных клеток и неподвижной линзой,— неэффективный орган зрения. Разрешающая способность фасеточного глаза усилена путем увеличения числа фасеток в сотни и тысячи раз. Фасеточные глаза в целом дают мозаичное изображение предметов по их мельчайшим движениям, но без конкретизации деталей.
По иному пути пошла эволюция органа зрения в группах, где происходило образование глазного пузыря с жидкостью и подвижного хрусталика — линзы. Эти изменения способствовали собиранию лучей позади линзы и тем самым повышению чувствительности глаза к свету. Разрешающая способность глаза еще более усилилась с образованием в нем сетчатки — группы светочувствительных клеток. Такие глаза характерны для головоногих моллюсков (кальмаров и осьминогов).
Дальнейшая эволюция привела к еще большему увеличению возможности восприятия глазом световых импульсов. Этому способствовали увеличение кривизны хрусталика, возникновение соответствия между главным фокусным расстоянием и расстоянием от хрусталика до сетчатки, развитие зрачка, глазной мускулатуры, светочувствительных клеток типа колбочек и палочек и т.д.
Выше были описаны этапы филогенетического развития органов зрения. Данные современной генетики показывают, что все без исключения изученные в этом отношении признаки и свойства органов зрения оказываются подверженными мутационной изменчивости (описано, например, несколько десятков мутаций глаз у дрозофил). Доказательством развития глаз под влиянием естественного отбора служат упрощение его строения и даже редукция (у пещерных животных — троглобионтов, у обитателей мутных потоков и др.). В этих случаях происходит быстрое накопление дезинтегрирующих мутаций, приводящих к разрушению органа зрения (прежде они устранялись стабилизирующей формой отбора).
Физиологические адаптации. Многочисленны примеры физиологических (функциональных) адаптаций. Например, для растений известен комплекс физиолого-биохимических мутаций, приведших к выработке приспособлений, которые связаны с устранением недостатка кислорода: использование кислорода фотосинтеза, нитратов и органических кислот как дыхательного материала, переключение путей метаболизма и т. д. Разнообразны и физиологические механизмы приспособления растений и животных к недостатку воды или неблагоприятным температурам. Отбор во всех этих случаях способствовал выживанию и преимущественному размножению индивидуумов — носителей благоприятных мутаций в соответствующих условиях.
Процессы фотосинтеза представляют пример закрепления в эволюции комплекса сложных приспособлений на структурном и биохимическом уровнях. В этом комплексе выделяются следующие этапы: синтез одного органического вещества за счет другого, смена использования состава веществ в качестве доноров электронов для восстановления CO2 (фотоорганотрофия: донор — органическое вещество, фотолитотрофия: донор — неорганическое вещество, фотогидротрофия: донор — вода). Соответственно в этом ряду выделяются следующие группы организмов: гетеротрофы — облигатные гетеротрофы — факультативные фотогетеротрофы — облигатные фотогетеротрофы — облигатные фотоавтотрофы — фоторедукторы — фотосинтетики.
Любой из перечисленных этапов метаболизма состоит из сложного комплекса реакций, каждая из которых подвержена мутационным изменениям (многие такие мутации хорошо изучены: так, по синтезу хлорофилла описано ныне более 200 только ядерных мутаций).
Общим для адаптаций в живой природе является механизм возникновения: во всех случаях достаточно подробного анализа оказывалось, что приспособление возникает не сразу в готовом виде, а длительно формируется в процессе эволюции. Это не оставляет сомнения в том, что приспособления любого масштаба всегда возникают в природе в процессе естественного отбора в пределах вида.
После описания примеров адаптаций сформулируем определение понятия «адаптация» и покажем механизм возникновения адаптаций.
