«Обезвреживание» мутаций в эволюции. Значительная часть вновь возникающих мутаций биологически «хуже» исходной нормы. Такое положение не удивительно. Нормальный комплекс генотипов природной популяции — результат длительного отбора лучших комбинаций, поэтому большинство отклонений от нормы должны оказаться «вредными» уже потому, что случайное изменение сложного механизма не может его улучшить. В то же время безграничная изменчивость очень легко может разрушить сложнейшие системы взаимодействия, начиная от взаимодействия молекул внутри клетки и кончая взаимодействием особей в популяциях и популяций в биогеоценозах.
Это противоречие решается возникновением в эволюции полового процесса, ведущего к обезвреживанию мутаций путем перевода их в гетерозиготное состояние.
Известно, что многие мутации в гетерозиготном состоянии не только не понижают, а часто повышают относительную жизнеспособность особей. Механизм, поддерживающий гетерозиготность особей, связан с возникновением полового размножения. Половой процесс в целом (включая явление гетерозиготности), с одной стороны, позволяет сохранять в популяциях вновь возникающие мутации, которые служат необходимым материалом для дальнейшего протекания процесса эволюции. С другой стороны, половой процесс обезвреживает последствия возникновения мутаций, «скрывая» их в гетерозиготном состоянии. Поэтому можно сказать, что возникновение разнополовости — одно из крупнейших эволюционных приобретений.
Еще сейчас существуют живые организмы, не имеющие типичного полового процесса (бактерии, цианеи). У них только один набор генов, и поэтому каждая новая мутация сразу же проявляется в фенотипе. В борьбе за существование позволяет им выжить на протяжении миллиардов лет только высокий темп размножения. Образно говоря, они «не помнят прошлого и не готовятся к будущему», не имея запасов генетической изменчивости, скрытой в гетерозиготном состоянии. Высокая численность и способность к размножению позволяют им приспосабливаться во всех встречающихся на планете условиях существования за счет выживания лишь немногих мутантных особей.
Сказанное касалось тех мутаций, которые при своем возникновении оказываются в той или иной степени вредными для особи. Таких мутаций большинство, но какой-то небольшой процент вновь возникающих с самого начала может принести в конкретных условиях не вред, а пользу для особи. Как бы ни была мала доля таких мутаций, они в грандиозных временных масштабах процесса эволюции сами по себе смогут сыграть заметную положительную роль. Примем, что одна подобная мутация возникает на один миллиард обычных; частота возникновения мутации по данному локусу составит 1 на 100 000 гамет (в природе наблюдается частота от 10—4 до 10—9 ); общее число локусов составляет 10 000 (у высших организмов — много больше); численность вида в одном поколении составляет 100 млн особей (численность майских жуков только на 1 км2 может достигать 1 млн особей); каждая особь производит за жизнь 1000 гамет (число сперматозоидов, продуцируемых человеком за период половой зрелости, составляет несколько биллионов, число яйцеклеток — около 400).
При этих исходных условиях в каждом поколении возникнет по всем локусам у всего населения такого гипотетического вида 10 млрд мутаций, из которых только 10 000 будут полезными. Но если учесть, что среднее время существования вида составляет несколько десятков тысяч поколений (см. гл. 12, 13), то за время существования вида в его недрах возникнет около 10 млн мутаций, которые могут быть подхвачены отбором.
При эволюционной оценке мутаций необходимо учитывать и различный характер проявления мутаций в разных условиях. Мутации, вредные в одних условиях, могут оказаться (и оказываются) полезными в измененных условиях (табл. 9.1).
| Таблица 9.1. Жизнеспособность особей Drosophila funebris — носителей разных мутаций — по отношению к жизнеспособности особей «дикого типа» того же вида, % (по H.B. Тимофееву-Ресовскому, 1934) | |||
| Мутация | Жизнеспособность при температуре, °C | ||
| 15—16 | 24—25 | 28—30 | |
| eversae | 98,3 | 104,0 | 98,5 |
| abnormal abdomen | 96,2 | 88,0 | 80,7 |
| miniature | 91,3 | 69,0 | 63,7 |
| bobbed | 75,3 | 85,1 | 93,7 |
Приведенные в таблице данные показывают, что при 24—25°С (диапазон температур, отличный от нормального, составляющего у этого вида около 20°С) носители мутаций eversae у Drosophila funebris оказываются более жизнеспособными, чем исходные природные особи. Естественно предположить, что при некотором потеплении в природе мутантные особи будут получать заметное преимущество перед исходной формой и быстро распространяться в природе, используя ранее малодоступные для вида условия существования.
В процессе возникновения нелетающих форм насекомых на океанических островах (этот случай гипотетически был рассмотрен еще Ч. Дарвином) даже более резкие мутации этого типа, вызывавшие редукцию крыльев, оказывались полезными, так как особи с развитыми крыльями сносились ветром в море и погибали в большем числе.
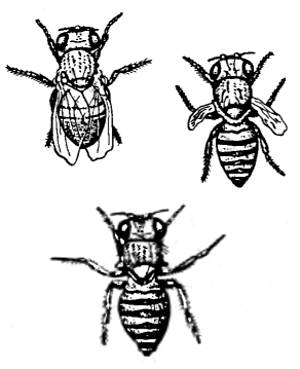
Рис. 9.2. Внешний вид трех мутантных форм дрозофил с редуцированными крыльями. См. текст (из А.С. Северцова, 1980)
У классического объекта генетиков дрозофилы описано немало мутаций, затрагивающих крылья, вплоть до их полной редукции. На рисунке три из них: cut («обрезанные крылья»), vestigial («зачаточные») и apterous («бескрылость») (рис. 9.2). Французский генетик Ж. Тесье содержал экспериментальную популяцию дрозофил, в которой мутанты vestigial были смешаны с диким типом в открытых ящиках на террасе рядом с морем. Через два месяца численность мух с зачаточными крыльями поднялась с 2,5 до 67%. В безветренные дни число гомозигот vestigial не повышалось, а когда ящики с популяцией перенесли в закрытое от ветра помещение, «дикие» мухи, как более плодовитые, стали вытеснять бескрылых. Появление бескрылых насекомых на океанических островах в результате естественного отбора подтвердилось экспериментально.
Говоря о вредности тех или иных мутаций, мы до сих пор рассматривали каждую отдельно взятую мутацию порознь. Однако в природе мутации встречаются всегда в комбинациях друг с другом. Эффект комбинирования может быть положительным для особи в целом, повышая ее жизнеспособность (табл. 9.2). Эта особенность мутаций хорошо известна в практике животноводства и растениеводства: часто две мутантные формы, маложизнеспособные порознь, объединенные в потомстве, обладают ценными хозяйственными свойствами.
Из данных табл. 9.2 видно, что в одном случае (eversae X singed) жизнеспособность комбинации мутаций оказалась выше исходной для «дикого типа», а в другом (miniature × bobbed) — выше жизнеспособности обеих исходных форм.
| Таблица 9.2. Жизнеспособность особей Drosophila funebris — носителей комбинаций двух мутаций, % от жизнеспособности «дикого типа» того же вида (по Н.В. Тимофееву-Ресовскому, 1934) | |
| Комбинации мутаций | Жизнеспособность при 24—25°C |
| eversae×singed | 103,1 — + |
| miniature bobbed | 96,6 + + |
| eversae×bobbed | 85,5 — + |
| singed×abnormal abdomen | 76,6 — — |
| singed×miniature | 67,1 — — |
* «— —» — Пониженная жизнеспособность сравнительно с обеими исходными мутациями; «+ +» — повышенная жизнеспособность сравнительно с обеими исходными мутациями; «— +» — пониженная жизнеспособность сравнительно с одной исходной формой и повышенная по отношению к другой.
Обобщая данные о вредности и полезности мутаций в процессе эволюции, можно сказать, что в природе мутации необходимы для нормального существования популяции в изменяющихся условиях и для освоения новых, ранее недоступных условий существования. Вредность же мутаций в основном нейтрализуется переводом их в гетерозиготное состояние.
