Половой состав популяции. Известно, что генетический механизм определения пола обеспечивает расщепление потомства по полу в отношении 1:1 (первичное соотношение полов). В силу неодинаковой жизнеспособности мужского и женского организма (разная жизнеспособность — эволюционно выработанный признак) это первичное соотношение порой уже заметно отличается от вторичного (характерно при родах у млекопитающих) и тем более заметно отличается от третичного, характерного для взрослых особей. У человека, например, вторичное соотношение полов составляет 100 девочек на 106 мальчиков, к 16—18 годам это соотношение (в результате повышенной мужской смертности) выравнивается, к 50 годам составляет 85 мужчин на 100 женщин, а к 80 годам — 50 мужчин на 100 женщин.
И у животных, и у растений вторичное и третичное соотношение полов может колебаться в значительных пределах у разных видов. У некоторых насекомых встречаются популяции, состоящие практически из одних самок в результате сложных генетических процессов, влияющих на элиминацию самцов. Только из самок состоят популяции ряда партеногенетических видов насекомых и ряда других животных. Интересно заметить, что доля партеногенетических самок в разных популяциях может значительно варьировать. Другой крайний случай в соотношении полов связан с эволюционно возникшей исключительно лабильной системой определения пола. Так, у корнеплода Arisaema japonica решающий фактор во вторичном определении пола — это масса клубней: самые крупные и хорошо развитые клубни дают растения с женскими цветками, а мелкие и слабые — с мужскими цветками. Личинка червя Bonellia viridis развивается в самку, если после некоторого периода жизни в море она не сможет прикрепиться к другой взрослой самке, если же личинка прикрепится к взрослой самке, то из нее всегда получается самец; самцы фактически паразитируют на самках, они в десятки раз меньше их по размерам и выполняют единственную функцию — оплодотворение яиц.
Знание экологической структуры популяции (включающей величину популяции и ее динамику в пространстве и во времени, возрастной и половой состав особей и другие особенности, изучаемые популяционной экологией) — обязательное условие глубокого изучения популяций как единиц эволюции в природе.
Основные морфофизиологические характеристики популяции. Поскольку внутри вида популяции обмениваются генетическим материалом, то популяции различаются количественным соотношением разных аллелей и частотами встречаемости того или иного фенотипа, т. е. статистически. Однако не всегда удается сразу найти такой признак, присутствие или отсутствие которого в заметной концентрации характеризует ту или иную популяцию по сравнению с соседними. В этих целях проще и надежнее сравнивать популяции по многим признакам. Особенно успешным такой подход бывает при использовании дискретных, качественных признаков, которые иногда называют фенами (А.В. Яблоков) или признаками-маркёрами генотипического состава популяции. Примерами фенов могут служить красная и черная окраска элитр у двухточечной божьей коровки, наличие или отсутствие пленки на зерне, зазубренная или гладкая ость у пшеницы, наличие и расположение отдельных мелких отверстий в черепе млекопитающих, форма отдельных костей черепа и швов между костями и т. д. Все подобные признаки отличаются качественностью, альтернативностью. Их легко учитывать в любой изучаемой совокупности особей в терминах частот встречаемости.
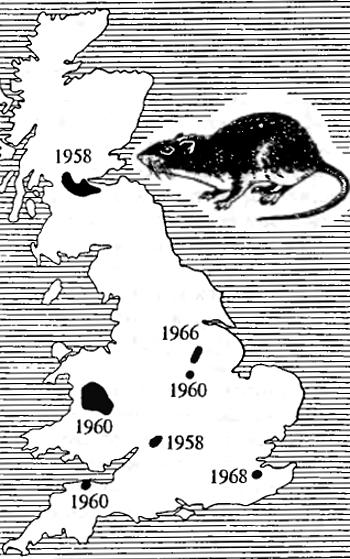
Рис. 7.2. Пример характеристики популяции по одному фену: места обнаружения устойчивых к антикоагулянту варфарину серых крыс (Rattus norvegicus) в Англии в разные годы (по Д. Друммонду, 1970)
В удачных случаях возможно характеризовать популяцию по отдельным, хорошо учитываемым признакам. На рис. 7.2 приведена карта встречаемости в Англии серых крыс (Rattus norvegicus), нечувствительных к сильному антикоагулянту варфарину, используемому для борьбы с мелкими грызунами. Генетическая природа этого признака сложна, так что нельзя сказать, что устойчивость к варфарину обусловлена действием какого-то определенного гена. Легкость обнаружения сделала этот физиологический признак удобным признаком-маркёром для определения границ, занимавшихся определенными популяциями до того периода, когда под влиянием бессознательного отбора нечувствительные к варфарину крысы распространились по всей Англии.
Сопоставление аналогичных данных, полученных по другим признакам, позволяет с большей степенью надежности характеризовать разные популяции, оконтуривать их границы, выяснять степень сходства с соседними популяциями, строить обоснованные гипотезы о путях исторического развития отдельных популяций и их групп, выяснять действие различных эволюционных факторов. В этих случаях приходится морфологические и физиологические признаки описывать, как и при изучении генетики популяции, в частотах встречаемости, т. е. подходить к изучению популяций с генетических позиций.
Фенетический подход не исключает традиционных способов сравнения популяций по размерам, массе, пропорциям тела составляющих их особей. Следует лишь подчеркнуть, что получаемые традиционными методами результаты крайне трудно использовать для точного эволюционно-генетического сравнения популяций из-за значительной полигенности и крайне малой наследуемости практически всех размерно-весовых характеристик особей.
