Корни пунктуализма уходят и в доктрину де Фриза о кратких периодах мутаций, когда, подобно взрыву, рождается множество новых видов, чтобы затем немногие из них, выдержавшие испытание на жизнеспособность, надолго остановились в своем развитии.
Через несколько лет Гулд и Элдридж (Gould, Eldredge, 1977) опубликовали новый, существенно видоизмененный вариант гипотезы. Если в первой публикации они ограничились описанием характера процесса видообразования, то теперь расширили рамки своей концепции, включив в нее гипотезу отбора видов Стэнли (Stanley, 1975), и заявили, что в этом новом виде их концепция должна стать основой целостной теории макроэволюции.
Как неоднократно отмечали Гулд и Элдридж, внутри популяционные дифференцировки, описываемые классической популяционной генетикой, имеют к видообразованию и макроэволюции очень малое отношение. Глубина и быстрота морфологических преобразований, связанных с последними, зависят главным образом от изменений регуляторных систем генома. Этим положением перечеркивалась идея Симпсона о необходимости синтеза данных палеонтологии с генетикой популяций и возрождался путь к альянсу с биохимической генетикой, провозглашенный Шиндевольфом.
В статье приводился новый палеонтологический материал, относящийся к радиоляриям, аммонитам, трилобитам и отчасти гоминидам, и подчеркивалась важность изучения эволюционного стазиса, темпов эволюции, а также сопоставления данных палеонтологии с выводами наук о ныне живущих организмах.
Идея прерывистого равновесия возникла как альтернатива градуалистической модели эволюции, идеально воплощенной в теории Дарвина. В XX в. оплотом градуализма традиционно оставались классическая популяционная генетика и, соответственно, синтетическая теория. На позициях градуализма до сравнительно недавнего времени стояли и молодые палеонтологи.
Градуализм органически связан с представлением об эволюции как филетическом процессе. На первый взгляд может показаться, что подобное представление противоречит дарвиновской схеме дивергентного формообразования. Вспомним, однако, что у Дарвина промежуточные формы вымирают, а крайние сохраняются, продолжая эволюционировать в направлении однажды начавшейся изменчивости. В работах Райта 1930-х годов доказывалось математически, что наиболее благоприятные условия для успешной работы естественного отбора создаются в больших сложных популяциях, а это равнозначно утверждению, что филетическая эволюция является наиболее быстрой. Надо сказать, что важной опорой градуализма была трактовка эволюции исключительно как процесса прогрессирующего адаптациогенеза: если первичные полезные изменения очень малы, значит, весь процесс видообразования складывается из последовательной аккумуляции огромного их числа.
Против этих особенностей градуалистической концепции как раз и выступили создатели «прерывистой» модели. Весь пафос их критики сосредоточился на доказательстве того, что, если бы эволюция шла исключительно путем филетического градуализма, органический мир современной эпохи, вероятно, не поднялся бы в своем развитии выше уровня палеозойских организмов. Имеющийся фактический материал свидетельствует, что филетическая эволюция реально существует, но происходит крайне медленно. Прямые наблюдения (Ehrlich, Raven, 1969; Endler, 1973, 1977; Stanley, 1979 и др.) подтверждают факт весьма слабого обмена между внутривидовыми популяциями многих ныне живущих видов и, соответственно, очень медленное распространение в них удачной комбинации генов, что и препятствует быстрой филетической эволюции. А без обмена генами объяснить такую эволюцию можно было бы только с помощью допущения, что параллельные (однонаправленные) и быстрые генетические изменения претерпевают все субпопуляции вида. Ныне показано, что в кризисные для экосистем моменты истории дело обстоит именно так, но в 70-е годы до такого понимания эволюционного преобразования наука еще не доходила.
Пунктуалисты противопоставляют филетическому градуализму модель прерывистого квантового видообразования, утверждая, что видообразовательный, или кладистический, процесс составляет большую часть содержания эволюции. Самое оригинальное в этой модели – всегда множественное видообразование, дающее целую гамму разных видов, из которых только один выживает и будет существовать в неизменном состоянии в течение миллионов лет – до нового видообразовательного события. Схематично прерывистое видообразование изображается в виде линий, ветвящихся под прямыми углами, почему гипотезу прерывистого равновесия называют еще «прямоугольной (rectangular) эволюцией» (рис. 22).
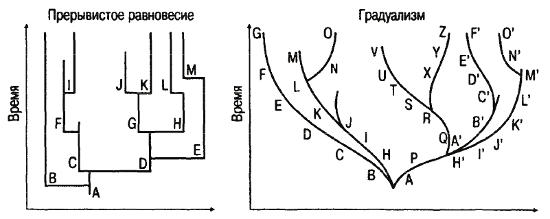
Рис. 22. Графическое изображение моделей прерывистого равновесия и градуалистического видообразования (Майр, 1982)
Полемика между сторонниками модели «экстраполяционного адаптивного филетического градуализма» и приверженцами пунктуализма (Gould, 1982а. Р. 137) в течение 10–15 лет заполняла страницы многих периодических изданий, неизменно возни-кала на международных научных форумах и пока еще не привела к окончательному результату. Градуализм или пунктуализм? Этот вопрос все еще дискутируется в современном эволюционном учении (Колчинский, 2002).
Для того чтобы дать более полное представление об обеих моделях, мы приводим их основные положения (табл. 2).
| Таблица 2. Сравнение гипотез постепенного и прерывистого видообразования | ||
| Фактор | Постепенное видообразование | Прерывистое видообразование |
| Источник изменчивости | Главным образом точковые мутации структурных генов | Мутации регуляторных генов и макромутации |
| Дивергенция | Медленная, непрерывная, пропорциональная числу поколений | Быстрая и непропорциональная числу поколений |
| Изоляция | Вторична; обычно аллопатическая | Чаще первична |
| Отбор | Внутри видовых популяций | Между видовыми популяциями |
| Адаптации | Очевидные и совершенные | Не столь очевидные и менее совершенные |
Следует, конечно, иметь в виду, что вопрос о соотношении прерывистой (быстрой) и постепенной (медленной) эволюции прежде всего эмпирический, и попытки его решения с помощью косвенных соображений и теоретических спекуляций вряд ли можно признать правомерными. Приоритетное значение должен иметь поэтому скрупулезный анализ палеонтологического материала. Для доказательства того, что эволюция той или иной группы была прерывистой, необходимо располагать непрерывными палеонтологическими сериями за длительный отрезок геологического времени, которые позволили бы зафиксировать периоды как стазиса, так и быстрой видообразовательной эволюции. Нужно также иметь возможность точно датировать (обязательно по абсолютной шкале) короткие интервалы в пределах данной последовательности.
Все эти условия были соблюдены в подробном исследовании кайнозойских брюхоногих и двустворчатых моллюсков из мощной толщи пресноводных отложений озера Туркана (Кения), осуществленном П. Уильямсоном (Williamson, 1981). Приблизительно из ста слоев осадочных пород, прослоенных вулканическими туфами точно установленного возраста, им был добыт и биометрически изучен массовый материал (около 3700 раковин), относящийся к 13 филумам. Благодаря этому Уильямсон имел возможность достоверно документировать все переходы в изменении морфологических признаков.
В итоге оказалось, что после периода стазиса, длящегося в течение 2-3 млн лет, новые виды возникают в интервале от 5 до 50 тыс. лет. Для медленно эволюционирующих моллюсков это достаточно короткий срок. Уильямсон констатировал, что ни в одной из изученных линий не было отмечено постепенных переходов. В моменты стрессовых ситуаций, создававшихся периодическими пересыханиями озер, у моллюсков наблюдалась сильная морфологическая изменчивость, на базе которой быстро формировались новые виды.
