Одним из первых, кто воспринял в Советском Союзе концепцию Берга после, казалось, ее полного забвения, был известный палеонтолог Б.Л. Личков (1965), горячий сторонник Ж. Кювье и автор одной из сопряженных с геологическими циклами гипотезы эволюции. По его убеждению, все прогрессивное развитие органического мира совершалось не на основе случайностей, а в силу строгих закономерностей. Однако уточнять эти закономерности, а тем более раскрывать их природу Личков не стал.
Эту миссию взяли на себя талантливый палеоботаник и теоретик-эволюционист, рано ушедший из жизни, СВ. Мейен и тесно сотрудничавший с ним Ю.В. Чайковский. Можно сказать, что они стали главными продолжателями Берга и некоторым образом Любищева в силу своей естественной склонности искать закон и порядок там, где его особенно трудно обнаружить. Мейен и Чайковский вообще стремились к «номотетизации» биологии, но, в отличие от господствовавшей тенденции к осуществлению этого намерения с помощью редукционистской методологии, они решали эту задачу путем познания биологического разнообразия.

Сергей Викторович Мейен (1935-1987)
Как известно, порядок в разнообразии, изучаемом типологией (у Чайковского – диатропикой), создает систематика (таксономия), которая опирается на данные о строении организмов, или, точнее, об их признаках. Эту науку о признаках Мейен (1977) предложил называть мерономией. В отличие от таксономии, распределяющий формы организмов по группам, в случае мерономии мы делим организм на части – по морфологическим, физиологическим или экологическим признакам, а классифицируя эти последние, получаем мероны («классы частей»). Примерами меронов могут служить любые части целого (органы, ткани, определенный тип клетки, физиологическая функция и т.п.), общие для данного таксона. В сумме они составляют его архетип, или план строения. Мерономия обеспечивает таксономию «признаковым пространством и данными о соотношении признаков у разных объектов» (Мейен, 1978. С. 496).
Теперь, сравнивая ряды параллельных таксонов, Мейен констатировал, что у них наблюдается сходный, а иногда и идентичный набор меронов. Чем ближе друг другу таксоны, тем больше число одинаковых меронов. У видов одного рода они почти все совпадают. Эту повторяющуюся последовательность меронов в паралельных таксонах Мейен назвал рефреном (там же, с. 501).
По аналогии с периодическим законом в химии он считал полезным графическое отображение рефренов в виде таблицы, где по горизонтали представлены изменения мерона в рядах сравниваемых таксонов. Вертикальные столбцы означают тогда одинаковые состояния выбранного мерона в этих таксонах (аналог одинаковой валентности в таблице Менделеева). Фактически это те же гомологические ряды Вавилова, только сведенные в таблицу. Мейен считал, что подобная форма записи ценна не столько для систематики, сколько для понимания процесса эволюции.
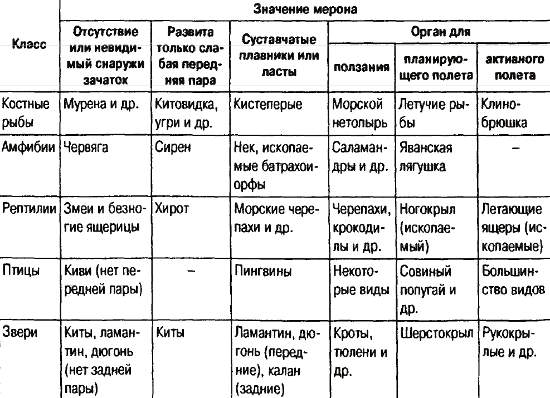
Рис. 19. Рефрен мерона «парные конечности» (из: Чайковский, 1990)
Так, при подобной записи мерона «парные конечности» (рис. 19) для всех классов позвоночных выявляется общий рефрен: от полного отсутствия обеих пар до образования органа полета – крыла. Таблица наглядно демонстрирует возникающее в силу закона параллелизмов признаковое пространство, позволяющее предсказывать, что может и чего не может быть в эволюции. В природе, правда, вовсе не обязательно должно существовать такое число разных форм, какое способно занять все клетки таблицы. Так, возвращаясь к мерону «парные конечности», надо отметить, что в истории никогда не было крылатых амфибий и птиц без задних конечностей. Мейен справедливо замечает, что без выявления всего мыслимого разнообразия нельзя установить и существования многих запретов.
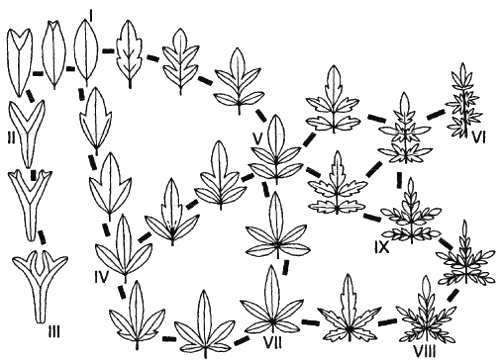
Рис. 20. Типы расчленений листовой пластинки (из: Мейен, 1973)
Один из самых наглядных примеров рефрена на ботаническом материале – типы расчленения листовой пластинки (рис. 20). Хотя в типологии обычен поиск закономерности в повторяемости признаков внутри таксонов, самая суть рефренов, по мнению Мейена, в выявлении закономерностей в изменчивости признаков между таксонами. Данные о них фрагментарны. Но без знания рефренов заполнить по отдельности все разновидности переходов между меронами – столь же невыполнимая задача, как заполнить все склонения каждою существительного, если не знать правил склонения.
Нам представляется весьма важной также еще одна закономерность формообразования в эволюции, выделенная Мейеном и названная им транзитивным полиморфизмом (Мейен, 1978). Она состоит в том, что новый таксон рождается с тем же набором составляющих его таксономических единиц или форм, существовавших у предкового таксона, которые повторяют и соответствующие признаки (рефрен). Причем даже небольшое число уцелевших особей способны восстановить все внутренее разнообразие истребленного таксона. Иными словами, разнообразие порождает разнообразие, на каком бы уровне мы его ни рассматривали, и только что сказанное – его следствие. Существование подобной закономерности в сочетании с происходящей при смене поколений свободной комбинаторикой признаков чрезвычайно усложняет обнаружение таксонов-предков и восстановление филогении, а то и вовсе делает это занятие бессмысленным. Эволюция идет и сразу «заметает» за собой следы.
Надо полагать, что, исследуя эту закономерность, Мейен знал, что идея об эволюционном переходе внутривидовой структуры от вида к виду уже существовала (Алтухов, Рычков, 1972). Мейен только расширил ее рамки.
Ученый обратил внимание на то, что в формулировке закона Вавилова речь идет только о повторности признаков от таксона к таксону, но в ней ничего не говорится о повторности в правилах их преобразования. С точки же зрения выявления типологических закономерностей это имело бы особое значение. Гораздо важнее самих морфологических параллелизмов тот факт, что, например, в разных семействах цветковых наблюдаются одинаковые тенденции в преобразовании морфологических особенностей. Именно общность тенденции, наличие одного правила преобразования позволяют предсказывать и целенаправленно искать недостающие члены параллельных рядов.
Мейен не ограничился только упорядочением закономерностей в рядах изменчивости, установленных своими предшественниками; он идет дальше, расширяя рамки самого закона Вавилова. Он убедился, что полнота проявлений параллелизма не всегда связана с генетической общностью. Сплошь и рядом она, напротив, обратно пропорциональна систематической близости таксонов. Так, параллелизм между головоногими моллюсками и фораминиферами по спиралям раковины гораздо полнее, чем между головоногими и брюхоногими по тому же признаку; параллелизм жизненных форм кактусов и молочаев полнее, чем между кактусами и более близкими к ним Caryophyllaceae. Немало примеров параллелизма в строении цветков и соцветий, листьев и филлодиев, в биохимических и генетических характеристиках далеких форм. Еще более удивительны параллелизмы между живыми и неживыми объектами, где уже не приходится говорить о конвергенции ввиду сходных условий существования. Примеры таких параллелизмов собрал еще д'Арси Томпсон (1942), а ближе к нашему времени – ЮЛ. Урманцев (1970, 1988) и А. Лима-де-Фариа (1991). Урманцев, в частности, подтвердил сходства гомологических рядов в развитии животных и растений с таковыми спиртов и углеводородов, установленные, соответственно, Э. Копом и Н.И. Вавиловым; обнаружил сходство между 9 изомерами венчика барбариса и 9 изомерами инозита, сходство генома с языком (речью), эволюционной генетики со сравнительным языкознанием и многие другие. Объяснение существованию подобного рода параллелизмов Мейен, вслед за Урманцевым, усматривает в системной природе объектов (Мейен, 1975).
Названные случаи изоморфического сходства, не сводимые ни к генетической общности, ни к подобию условий существования, привели Мейена к выводу о существовании чисто морфологических (типологических) и нестатистических законов, являющихся наиболее общими и пока еще очень слабо изученными. В свете такого взгляда закон гомологических рядов Вавилова, очевидно, становится его частным проявлением.
Следует отметить еще один новый момент в трактовке все того же закона. Обычно не учитывается, замечает Мейен, что признаки радикала тоже иногда испытывают изменчивость, которая в силу своей редкости считается тератологической. Эта изменчивость тоже следует закону Вавилова, причем часто признак, расцениваемый как уродство в одном таксоне, становится нормой в другом. Этот частный случай, описанный Н.П. Кренке (1933-1935), Мейен предложил называть правилом Кренке.
Среди ученых – экспериментаторов и теоретиков – безусловно существуют в относительно чистом виде две категории: «примирители», склонные к компромиссу и объединению кажущихся противоположностей в высшем синтезе, и «непримиримые», производящие выбор среди противоположностей и стремящиеся подавить бракуемую. Любищев принадлежал ко второй, а Мейен – к первой. Но на путях к синтезу Мейена постигла явная неудача.
Развивая стержневую идею номогенеза, он, в отличие от Берга, не создал целостной концепции. Берга интересовали и закономерности эволюции, и ее движущие силы. Собственно, с постулата, что автономический ортогенез – главная побудительная сила всякого прогрессивного развития, и начинается изложение его концепции о направленности эволюции. Во времена оные за это Берга наградили нелестными эпитетами – считали идеалистом, автогенетиком, преформистом, виталистом… Можно ли было в конце XX в. как-то развить, усовершенствовать это ядро номогенеза? Это нелегко. Для этого нужно предложить свое видение проблемы, так как в первую очередь всех интересуют именно факторы эволюции.
Мейен принял в качестве факторов эволюции мутации и отбор в том виде, как они еще существуют в СТЭ. Более того, он считал своей главной стратегической задачей поиски путей снятия противоречий между тихогенезом (селекционизмом) и номогенезом (Мейен, 1974, 1978, 1984а) и полагал, будто она достигнута с созданием общей теории систем, в рамках которой дивергенция и параллелизм стали дополнительными понятиями.
А между тем фундаментальное и в наши дни исключительно актуальное обобщение Мейена о тропиках как «колыбели» и «музее» растительного богатства планеты (Мейен, 1984б, 1986) находится в разительном противоречии с теорией селектогенеза. На огромном ископаемом материале ему удалось показать, что почти все наиболее крупные таксоны, включая семейства, возникли в фитохориях, лежащих в экваториальном поясе. И это стало возможно, констатирует Мейен, только благодаря тому, что естественный отбор здесь сильно «заторможен» и потому допускает всевозможные эксперименты природы.
Можно только гадать, как такой высокоэрудированный специалист с острым и проницательным умом мог закрывать глаза на очевидную несовместимость СТЭ и номогенеза.
