Дарвин не располагал прямыми доказательствами выживания наиболее приспособленных и конструировал воображаемые примеры. Один из них касается нектара растений и питающихся им насекомых. Представим себе, писал Дарвин, что нектар стал выделяться некоторыми растениями не на поверхности, а внутри цветков. Тогда к телу насекомых, залезающих в цветок ради этого сока, пристанет пыльца и будет затем перенесена на другие цветки и другие растения. Произойдет перекрестное опыление, которое обычно сообщает потомству большую силу и плодовитость и, следовательно, повышает его шанс на выживание в борьбе за существование. Если отбор в направлении все большего погружения нектарников в глубь цветка продолжится и в следующих поколениях, то возникнет новая форма растения, приспособленная к постоянному опылению насекомыми.
Другой гипотетический пример описывает воображаемые последствия едва заметного изменения длины или формы хоботка пчел, которые позволяют отдельным особям-носителям этого признака добывать нектар из цветков, не доступный другим пчелам. Если особи, обладающие такими полезными изменениями, будут из поколения в поколение сохраняться, то они образуют местную расу, приспособленную к этим цветкам и несколько отличную от пчел соседних местностей.
Из самого определения естественного отбора как механизма, приводящего к выживанию наиболее приспособленных, следует, что этот механизм ведет к совершенствованию адаптации и сообщает всей эволюции строго адаптивный характер. Таким образом, естественный отбор дает одновременно материалистическое объяснение и формообразованию, и происхождению органической целесообразности. В этом видят вторую историческую заслугу Дарвина.
Главным материалом для действия естественного отбора служат преимущественно мелкие, неопределенные наследственные изменения. Благодаря отбору из общего их числа сохраняются только полезные. Они суммируются им, накапливаются, усиливаются и становятся характеристиками все более крупных систематических категорий. Дарвин рисует, таким образом, эволюцию как преимущественно градуалистический процесс.
Достаточность едва уловимой изменчивости и постепенность селективного процесса великолепно передана Дарвином в следующих словах: «Выражаясь метафорически, можно сказать, что естественный отбор ежедневно и ежечасно расследует по всему свету мельчайшие изменения, отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно и невидимо, где бы и когда бы ни представился к тому случай, над усовершенствованием каждого органического существа в связи с условиями его жизни… Мы совершенно не замечаем этих медленно совершающихся изменений в их движении вперед, пока рука времени не отметит истекших веков…» (с. 330). Или еще одно прямое свидетельство пред-почтения градуализма: «Не может быть сомнения, что многие виды образовались крайне постепенно» (с. 456).
Естественный отбор в глазах Дарвина – творческая сила гораздо более могущественная, чем человек. Это признание рождает в душе Дарвина особый пафос. Он называет все усилия человека (значит, включая и плоды селекции) «жалкими» «в сравнении с теми, которые накопила Природа на протяжении целых геологических периодов». Естественный отбор – это сила, «столь же неизмеримо превосходящая слабые усилия человека, как произведения Природы превосходят произведения Искусства» (с. 315).
Дарвин специально останавливается на феномене полового отбора – склонности самцов или самок многих видов осуществлять селективное спаривание посредством прямой физической борьбы с соперниками или демонстрации наиболее привлекательных брачного наряда и поведения. В обоих случаях спаривание с более сильным или красивым партнером способствует появлению такого же потомства. Половой отбор совпадает, таким образом, по своему направлению с действием естественного отбора и рассматривается Дарвином как частная форма внутривидового отбора.
В деятельности естественного отбора есть одна важная закономерность, направленная на ослабление конкуренции. Она связана со свойством любого местообитания вмещать тем большее число особей вида, чем разнообразнее их потребности. В силу этого естественный отбор, осуществляя свою элиминирующую функцию, в числе менее приспособленных уничтожает и формы, промежуточные по своим потребностям между крайними вариантами. Тем самым крайние, наиболее уклонившиеся формы будут в меньшей степени конкурировать между собой за пищу, убежища, места размножения и в результате смогут увеличить свою численность и расширить ареал. Благодаря полезности такого процесса для вида различия между его уклоняющимися особями с каждым новым поколением будут возрастать, пока в конце концов эти группы особей не превратятся в самостоятельные разновидности, а затем – в новые виды. Процесс расхождения признаков, ведущий к формообразованию, Дарвин назвал дивергенцией и наглядно изобразил его в виде известной диаграммы.
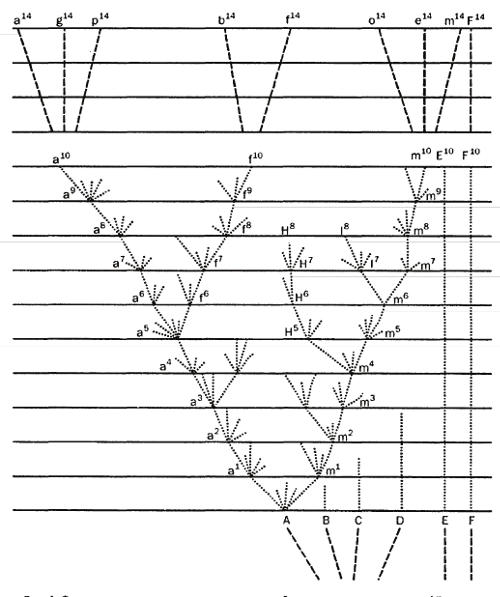
Рис. 1. Фрагмент диаграммы, иллюстрирующий расхождение признаков (Дарвин, 1939)
Воспроизводя самый важный фрагмент этой диаграммы (рис. 1), напомним, что на ней заглавными буквами А, В, С и далее обозначены родоначальные виды, строчными буквами с цифровыми индексами a1, a2,…, a10, a14 или m1, m2,…, m10, m14 и другими – степени изменчивости потомков родоначальных видов и последовательные ступени формирования рас, разновидностей, подвидов и видов. Каждая горизонтальная линия отделяет промежутки времени, соответствующие тысяче поколений. На диаграмме процесс дивергенции от вида А завершается образованием через 14 тыс. поколений формированием видов, обозначенных буквами от a14 до m14. В совокупности с видами другой части диаграммы они образуют новый род.
Но процесс расхождения признаков достижением этого этапа не завершится. Народившиеся виды еще слишком сходны между собой в основных потребностях и чертах морфологической организации. Чтобы меньше конкурировать между собой и с максимальной полнотой использовать все жизненные ресурсы местности, нужно продолжать однажды начавшуюся дивергенцию. Тогда образовавшиеся виды таким же дивергентным путем дадут новые дочерные виды, которые в совокупности составят новый род; совокупность родственных родов составит семейство; совокупность семейств – отряд и т.д. Так, все поднимаясь и разветвляясь, вырастает великое древо многообразия жизни. Одновременно формы, промежуточные между расходящимися потомками, будут вымирать.
Для отображения этого процесса образования высших таксонов пригодна та же диаграмма, если принять, что промежутки между горизонтальными линиями соответствуют не тысяче, а миллиону и более поколений или что они изображают последовательные слои земной коры.
Такова классическая дарвиновская схема видообразования и происхождения всех высших систематических категорий. Одновременно она отражает также монофилетический принцип эволюции и нарастания биоразнообразия, в основе которого лежит представление о происхождении каждой крупной систематической группы от одного общего корня (предка).
Внутривидовая конкуренция, дивергенция признаков и происхождение родственных групп организмов от общего корня составляют три неразрывных логических устоях теории Дарвина и придают ей ту цельность и внутреннюю стройность, которые всегда завораживали эволюционистов. Ее можно было либо принять целиком, либо целиком же отвергнуть. Посмотрим теперь, как эта столь искусно построенная теория соотносится с некоторыми самыми существенными фактами.
Нам представляется очень странным, как такой великий и объективный натуралист, каким был Дарвин, мог пройти мимо столь универсальных явлений, как взаимопомощь внутри вида и симбиоз. Почему в «Происхождении видов» нет ни слова о стадах и стаях высших животных и о самом факте совместной жизни общественных насекомых? Лишь много позднее, в книге «Происхождение человека и половой отбор» (1871), Дарвин признал, что жизнь в тесном сообществе составляет самое условие выживания всех видов общественных насекомых. А ведь эти и многие другие постоянные скопления организмов одного вида самим фактом своего существования опровергают положение Дарвина об особой роли внутривидовой конкуренции.
Дарвин не мог не знать и о многочисленных примерах мутуалистического симбиоза, хотя сам термин «симбиоз» был введен в науку только в 1879 г. Хорошо известно, что члены симбиотических систем, нередко образованных несколькими видами, реально облегчают друг другу жизнь, ослабляя негативные влияния внешней среды. При эндо- и внутриклеточном симбиозе происходит даже объединение основных метаболических процессов.
Возможно, во времена Дарвина случаи симбиоза представлялись еще единичными и ими можно было пренебречь. Но в наши дни симбиоз предстал как поистине планетарный феномен. Установлено, что из общего числа видов животных и растений, оцениваемого разными авторами в 2 или 12,5 млн (Global biodiversity status.., 1992), от 1/3 до 1/2 участвуют в симбиотических отношениях и таким образом в значительной мере избегают взаимной конкуренции и борьбы. Эта статистика говорит сама за себя.
Известно, что симбиоз, хотя он и является продуктом и своего рода «надстройкой» эволюции, восходит к очень давним временам. Возможно, симбиотические отношения зародились уже на ранних этапах становления жизни на Земле. Во всяком случае симбиоз выступал самостоятельным фактором эволюции на всем протяжении фанерозоя.
Главная ошибка Дарвина состояла в том, что он построил свою теорию, исходя из одной трагической стороны жизни. Думается, что игнорирование Дарвином положительных внутривидовых отношений у организмов, совпадающее с невниманием к положительным последствиям корыстолюбивых стремлений в учении Смита, не случайно. И это склоняет нас к мысли о справедливости суждения некоторых историков дарвинизма о том, что Дарвин сначала создал теорию, а потом подбирал под нее факты (Ghiselin, 1969). Закономерно, что односторонний взгляд Дарвина на движущие силы эволюции как борьбу и гибель слабых, воспринятый политиками, привел в XX в. к трагическим социальным последствиям.
Завершая анализ «зрелой» теории Дарвина, следует прямо сказать, что ни одно из ее принципиальных положений не сохранило исходного эволюционного значения и не получило фактического подтверждения. Перенаселение оказалось мнимым; внутривидовая конкуренция и борьба устраняются в природе многочисленными регулирующими механизмами, и часто вместо них наблюдаются взаимопомощь и сотрудничество; естественный отбор предстает как достаточно грубый механизм, не способный забраковать даже особи с явно уродливой организацией. Стало известно, что жизнеспособность аномальных форм определяется не на фенотипическом, а на генетическом уровне. И, конечно, при подобном качестве отбор никак не может выполнять кумулирующую, созидательную функцию – быть творческим фактором. Наконец, если естественный отбор не в состоянии подхватить слабую (индивидуальную) изменчивость, то дивергентный характер эволюции и преобладание в ней монофилии ставятся под вопрос. Все это позволяет отнести дарвинизм к разряду теорий, имеющих хотя и огромное, но теперь уже историческое значение.
Но за Дарвином в числе его многих других заслуг одна сохранится навсегда как непреходящая – метод конструирования теории. Как мы надеемся показать в последующих главах, им воспользовались в XX в. как сторонники, так и противники дарвинизма. К этой стороне теории Дарвина мы сейчас и перейдем.
