Глава II
Клеточные мембраны
На основании электронно-микроскопических исследований стало ясно, что важным структурным элементом клеточной организации является мембрана. Она не только покрывает клетку снаружи, отделяя ее содержимое от внешней среды, но и органеллы клетки (митохондрии, эндоплазматический ретикулум, комплекс Гольджи, лизосомы) построены из мембран. Ядро также покрыто мембраной. Роль мембран очень разнообразна, они имеют отношение почти ко всем функциям клетки. С деятельностью мембраны связано протекание строго согласованных биохимических реакций, ведущих к распаду и синтезу веществ, избирательная клеточная проницаемость, электрическая поляризация клетки, двигательная, рецепторная функция и многие другие. Биологические мембраны являются объектом изучения не только биологов, но и химиков и физиков.
Строение и химический состав клеточных мембран
После химической фиксации под электронным микроскопом мембраны выглядят как трехслойные структуры толщиной 7,5 нм (рис. 14). Согласно традиционной модели Даниели—Давсона—Робертсона, средний, более светлый промежуток соответствует непрерывному липидному бислою, в котором гидрофильные группы липидов обращены наружу. Белки располагаются на полярной поверхности липидов и вступают с ними в электростатическое взаимодействие (рис. 14, А). Американский ученый Робертсон полагал, что подобная элементарная мембрана лежит в основе строения любой клеточной мембраны. Однако имеются данные, указывающие на то, что, несмотря на общее сходство в строении мембранных структур, имеются между ними существенные различия. В результате дальнейшего, более детального изучения мембранных структур в универсальную модель были введены некоторые усложнения. Кроме того, были предложены новые модели, в которых основная роль в поддержании структурной организации мембран приписывается гидрофобным связям. Согласно модели Ленарда и Зингера, полярные головки фосфолипидов вместе с ионными боковыми группами структурного белка расположены на поверхности мембраны, а неполярные цепи белка и липиды располагаются внутри мембраны и стабилизируются здесь гидрофобными взаимодействиями (рис. 14, Б). Геометрическая модель Вандеркой и Грина состоит из двух слоев глобулярных белков; пространство между глобулами заполнено фосфолипидными молекулами, полярные головки которых расположены на водной поверхности мембраны (рис. 14, В). В отличие от этой модели в жидкостно-мозаичной модели Зингера и Николсона глобулярные белки и фосфолипиды не являются фиксированной структурой, они более подвижны и свободно перемещаются в мембране (рис. 14, Г).
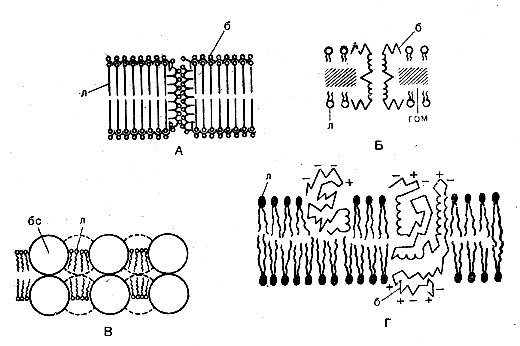
Рис. 14. Модели мембраны: А — по Даниели—Давсону—Робертсону; Б — по Ленарду и Зингеру; В — по Вандеркой и Грину, Г — по Зингеру и Николсону; б — белки, л— липиды; бс — белковые субъединицы; гомгидрофобная область мембраны
Таким образом, согласно новым моделям, срединная область мембраны не является непрерывным липидным слоем, а состоит из субъединиц — глобулярных белков, или липопротеинов.
Улучшение методов химической фиксации и применение физических (быстрое замораживание с последующей возгонкой витрифицированной воды в вакууме при низкой температуре) позволили выявить у многих мембран (митохондрий и эндоплазматического ретикулума) глобулярную структуру срединной гидрофобной области. Размер глобул составляет 5—9 нм. Глобулярность лучше выражена в тех мембранах, которые больше содержат белка. Большинство исследователей считает, что глобулы представляют собой липопротоиновые субъединицы, из которых построены мембраны. Субъединицы глобулярной формы и разной величины (4—12 нм) обнаружены и в фотосинтезирующих мембранах растительных клеток.
Помимо глобулярной структуры были обнаружены и пристеночные, более гидрофильные слои аморфного электронно-оптически плотного материала. Они наблюдаются и с внешней и с внутренней цитоплазматической стороны мембраны и достигают значительной толщины (до 0,1 мкм). Пристеночные слои богаты ферментами и мукополисахаридами и вместе с основной тонкой мембраной они составляют единый структурный и функциональный комплекс.
Благодаря методике дифференциального центрифугирования получают фракции разных мембран — поверхностных, митохондриальных, мембран эндоплазматической сети. Химический состав их оказался различным. Так, инертные миелиновые мембраны нервного волокна, выполняющие главным образом роль изолятора, в своем составе имеют 80% липидов. Более активные ядерные мембраны содержат 35% липидов, митохондриальные — 30%, протоплазматические мембраны скелетных мышечных волокон — 15%, а мембраны печеночных клеток — всего 10%, так что преобладающим компонентом в них является белок. Состав липидов у разных мембран отличается друг от друга (табл. 2).
| Таблица 2. Липидный состав мембран (%) некоторых клеток (Korn, 1966) | |||||
| Липиды | Миелин | Эритроциты | Митохондрии | Микросомы | Кишечная палочка |
| Холестерин | 25 | 25 | 5 | 6 | 0 |
| Фосфатидилэтаноламин | 14 | 20 | 28 | 17 | 100 |
| Фосфатидилсерин | 7 | 11 | 0 | 0 | 0 |
| Фосфатидилхолин | 11 | 23 | 48 | 64 | 0 |
| Фосфатидилинозитол | 0 | 2 | 8 | 11 | 0 |
| Фосфатидилгрицерол | 0 | 0 | 1 | 2 | 0 |
| Кардиолипин | 0 | 0 | 11 | 0 | 0 |
| Сфингомиелин | 6 | 18 | 0 | 0 | 0 |
| Цереброзид | 21 | 0 | 0 | 0 | 0 |
| Цереброзид сульфат | 4 | 0 | 0 | 0 | 0 |
| Церамид | 1 | 0 | 0 | 0 | 0 |
| Неидентифицированные | 12 | 2 | 0 | 0 | 0 |
Белковый состав также различен. Помимо белков и липидов мембраны содержат углеводы, неорганические ионы и воду.
Клеточные мембраны обладают асимметричной структурой и асимметричными функциональными свойствами. Так, строфантин оказывается более эффективным при действии его на внешнюю сторону мембраны, а кальций наиболее эффективен при действии на внутреннюю сторону. Обнаружена асимметрия в составе билипидного слоя: на эритроцитарных мембранах, на внешней половине билипидного слоя содержится больше холинсодержащих липидов, а на внутренней преобладают липиды с аминными группами. Асимметрия проявляется также в разной толщине внутреннего и внешнего мембранных слоев.
